陈红宇,张景,曾珍,杨娉娉,熊宇,张淼,周定安△
遗传性泛发性色素异常症(dyschromatosis universalis hereditaria,DUH)是一种罕见的遗传性皮肤病,此病由Ichikawa和Higara于1933年首次报道,主要表现为常染色体显性遗传,少数病例存在常染色体隐性遗传,该病典型临床表现为全身泛发无症状的色素沉着斑和色素减退斑[1]。在线人类孟德尔遗传(Online Mendelian Inheritance in Man,OMIM)数据库根据DUH 的连锁定位区域和致病基因不同分为DUH1(OMIM 127500)、DUH2(OMIM 612715)和DUH3(OMIM 615402)3 种类型。目前发现,在人类染色体6q24.2~6q25.2 区域的SASH1基因变异和染色体2q33.3~q36.1区域的ATP结合盒转运蛋白家族亚家族B 成员6(ABCB6)基因变异与DUH 发生相关。本研究拟探讨引起DUH 临床表型出现的SASH1基因变异是否影响黑色素合成。
1 对象与方法
1.1 研究对象
先证者,女,32岁,出生于中国湖北省,出生时皮肤正常,婴幼儿时期面部出现皮肤病变,青春期早期,患者颈部、肘部、膝关节和指骨关节开始出现色素沉着斑。成年后,面部、腹部、背部、手臂、大腿、小腿和臀部开始逐渐出现不规则形状、大小不一的色素沉着和色素减退斑,见图1。手掌、足底、口腔黏膜、头发、指甲和牙齿均正常。该患者经贵州医科大学附属医院和成都医学院2 名皮肤科专家确诊为DUH。本研究所涉及的患者一切资料均经贵州医科大学附属医院伦理委员会伦理审查(伦理号:2020073K),所有涉及研究对象均签署知情同意书。
1.2 主要试剂
小鼠黑色素瘤细胞B16、人皮肤黑色素瘤细胞SK-MEL-1、正常人皮肤黑色素细胞PIG1。SK-MEL-1 细胞的专用培养基购自于武汉普罗赛尔生命科技有限公司;Nano drop超微量分光光度计购自美国Thermo Fisher Scientific 公司;ABI 3730xl 测序仪购自美国应用生物系统公司;DNA 提取试剂盒,凝胶回收试剂盒购自天根生化科技(北京)有限公司;SASH1基因(NM_015278.5)的cDNA 序列由美国OriGene Technologies 公司合成;野生型pEGFP-C3 质粒购自美国Addgene公司;KOD-Plus定点突变试剂盒购自东洋纺(上海)生物科技有限公司;过表达野生型SASH1基因腺病毒(HAWT-SASH1-ADV)和过表达突变型SASH1基因腺病毒(HAT525R-SASH1-ADV)、聚凝胺由汉恒生物科技(上海)有限公司合成和提供;RPMI 1640 培养基、Dulbecco 改良Eagle 培养基、青霉素、链霉素和胰酶购自美国Invitrogen公司;胎牛血清购自维森特生物技术(南京)有限公司;RIPA 蛋白裂解液、Lipo8000™转染试剂、SignalUp™Western一抗稀释液、4%多聚甲醛固定液、免疫染色通透液(Triton X-100)、山羊血清购自上海碧云天生物技术有限公司;PVDF膜购自美国Promega公司;鼠抗人绿色荧光蛋白(green fluorescent protein,GFP)、血凝素(hemagglutinin,HA)、微管蛋白(β-tubulin)、甘油醛-3-磷酸脱氢酶(GAPDH)鼠抗人一抗购自艾比玛特医药科技(上海)有限公司;MITF一抗、Tyrosinase一抗购自英国Abcam公司;山羊抗兔IgG H&L(Alexa Fluor®594)、DAPI 和HRP 偶联的羊抗兔/鼠二抗购自英国Abcam 公司;兔抗人SASH1多克隆抗体购自美国Novus 公司;细胞黑色素含量比色法定量检测试剂盒购自上海杰美基因医药科技有限公司;限制性内切酶EcoRⅠ和BamHⅠ、T4 DNA连接酶购自美国NEB公司;大肠埃希菌Top 10 感受态细胞购自北京康为世纪生物科技有限公司。
1.3 方法
1.3.1 基因组DNA提取
采集先证者及其父母外周抗凝全血,使用DNA提取试剂盒提取3人DNA。Nano drop 超微量分光光度计测定DNA 样本浓度,DNA样本-20 ℃保存。
1.3.2 全外显子组测序及Sanger测序验证
将提取的3 人DNA 样本委托北京智因东方转化医学研究中心有限公司进行全外显子组测序。外显子组测序检出的SASH1c.1574C>G(p.T525R)突变位点,应用Premier 5.0软件设计该变异位点PCR引物:上游5'-AGCGGTCAAAGAGTG AGCAC-3' ,下游5'-CATGGAGCTCTGCCCACT-3' 。 ABI 3730xl测序仪进行Sanger测序,SeqMan软件对测序结果与标准序列进行比对。
1.3.3 生物信息学分析
应用PolyPhen-2(http://genetics.bwh.harvard.edu/pph2/index.shtml)软件预测SASH1-T525R 氨基酸替换对蛋白质结构和功能的影响。软件基于HumDiv 和HumVar 两个数据集的结果,给出一个分值,该分值范围为0~1,越接近1,说明氨基酸突变后对蛋白质结构和功能造成影响的概率越大。MEGA7软件对变异位点进行不同物种之间保守位点分析。
1.3.4 基因克隆、定点突变和病毒包装
Premier 5.0 软件设计SASH1基因全长引物,序列:上游5'-CCGGAATTCCGGATGGAGGACGCGGGAGCAGCTGGCCC GGGGCCGGAGC-3',下游5'-CGGGATCCCGCTACATGGCCT CAGGGCCTGGCGGCAGTTT-3'。以SASH1基因cDNA 为模板,利用全长引物和DNA 聚合酶进行PCR。PCR 扩增条件:95 ℃2 min;95 ℃30 s,65 ℃30 s,72 ℃1 min,共30 个循环;72 ℃5 min;4 ℃恒温。EcoRⅠ和BamHⅠ双酶切pEGFP-C3质粒和SASH1全长基因,酶切产物经连接、转化后获得pEGFP-SASH1-C3 重组质粒。挑取单个菌落于96 孔板中培养2 h,菌液PCR筛选阳性克隆。抽提阳性菌液质粒,用EcoRⅠ和BamHⅠ双酶切鉴定,并送测序验证插入序列的正确性。将已验证正确性的pEGFP-SASH1-C3质粒使用KOD-Plus诱变试剂盒将SASH1基因第1574位核苷酸C定点突变为G,构建pGFP-T525R-SASH1-C3 质粒,测序以验证该质粒的正确性。委托汉恒生物上海有限公司将野生型SASH1和突变型SASH1序列克隆到adeasy014-pAdEasy-EF1-MCS-CMVEGFP腺病毒载体中。
1.3.5 激光共聚焦显微镜明确SASH1在PIG1细胞的定位
将PIG1细胞以30%密度平铺在6孔板中,细胞培养24 h取出6 孔板,依次进行4%多聚甲醛室温固定30 min,0.2%Triton X-100冰上通透5 min,5%山羊血清封闭1 h,加兔一抗SASH1(1∶100)4 ℃过夜,次日用山羊抗兔IgG H&L 二抗避光1 h,漂洗后加DAPI 进行细胞核染色1 h,避光封片。染色后的片子用激光共聚焦显微镜扫描(蔡司LSM710),明确SASH1在PIG1 细胞的定位情况。其中对照组一抗用PBS 处理PIG1细胞,阳性组用SASH1单克隆抗体处理PIG1细胞。
1.3.6 细胞培养、转染、感染
B16 细胞培养在含有100 U/mL 青霉素和100 mg/L 链霉素、10%胎牛血清的RPMI 1640培养基中;SK-MEL-1细胞培养在SK-MEL-1专用培养基中;PIG1细胞培养在含10%胎牛血清的Dulbecco 改良Eagle 培养基中。所有细胞均在37 ℃、5%CO2培养箱中培养,每24 h 更换1 次培养基。将pEGFPSASH1-C3 和pGFP-T525R-SASH1-C3 质粒按照Lipo8000™转染试剂说明书转染B16 细胞48~72 h,不做任何处理的B16 细胞作为空白对照。利用聚凝胺将对照组、HA-WTSASH1组、HA-T525R-SASH1组腺病毒感染B16 细胞、SKMEL-1 细胞,按照制造商提供的程序进行,用免疫荧光显微镜观察感染效果。
1.3.7 Western blot 检测黑色素瘤细胞中SASH1、MITF、Tyrosinase蛋白表达
收集各组细胞,提取蛋白,4 ℃离心收集蛋白。每孔上样蛋白量为25µg,经12%SDS-PAGE,电泳后转至PVDF 膜,脱脂奶粉封闭。加一抗MITF(1∶400)、GFP(1∶400)、Tyrosinase(1∶500)、HA(1∶800)、GAPDH(1∶1 000)、β-tubulin(1∶1 000),4 ℃冰箱孵育过夜,次日二抗室温摇床孵育2 h后,采用化学发光仪进行显影。
1.3.8 体外黑色素定量T525R-SASH1是否影响黑色素合成
按照细胞黑色素含量比色法定量检测试剂盒的操作步骤将HA-WT-SASH1-ADV 和HA-T525R-SASH1-ADV 的B16 细胞和SK-MEL-1 细胞中的黑色素提取出来,通过光谱仪在405 nm 波长处测量吸光度(A),换算出黑色素含量,每组重复3次实验。
1.4 统计学方法
实验数据采用SPSS 16.0 软件进行数据分析,GraphPad Prism 5 进行绘图,计量数据采用均数±标准差(±s)表示,多组间比较采用单因素方差分析,组间多重比较采用LSD-t检验,P<0.05为差异有统计学意义。
2 结果
2.1 全外显子组测序结果
先证者存在SASH1c.1574C>G(p.Thr525Arg)的杂合变异位点,该变异位点位于人6号染色148 853 942 位置,美国医学遗传学与基因组学学会致病等级为致病性变异,千人基因组数据库、MAF 数据库目前未收录该变异位点;先证者父母该变异位点为野生型。测序结果提示先证者存在的该变异并非来自父母,而是新生突变,SASH1基因变异与先证者表型存在相关性。先证者SASH1基因染色体位于chr6:1488 53942,核酸改变c.1574(exon 14)C>G,氨基酸改变p.Thr525Arg(NM_015278),ACMG 致病等级为致病(PS2+PM1+PM2+PM),次要等位基因频率、单核苷酸多态性数据库、千人基因组和外显子组整合数据库均未收录。先证者父母该SASH1基因位点为野生型c.1574(exon 14)C。
2.2 Sanger测序验证
先证者存在SASH1基因c.1574C>G 杂合变异位点,其父亲和母亲未携带该杂合变异位点,见图2。
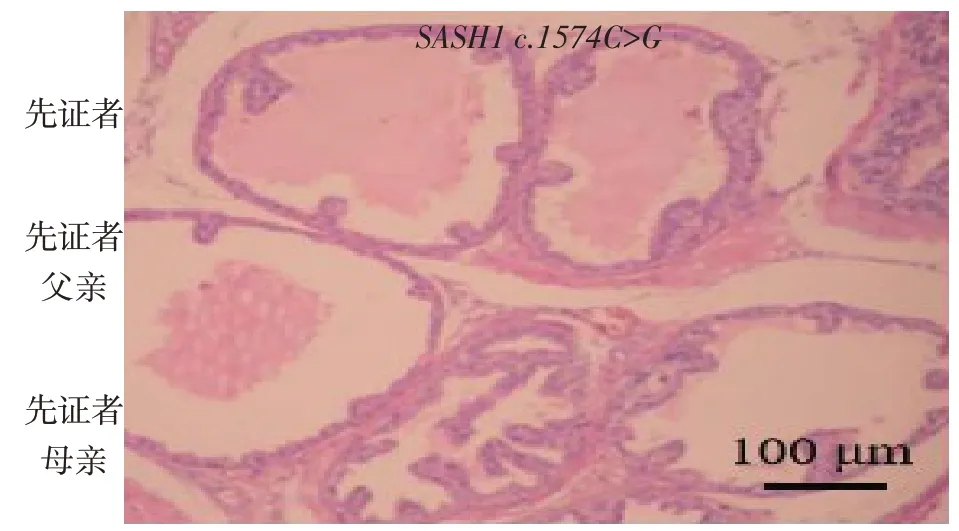
Fig.2 Sanger sequencing of SASH1 c.1574C>G of the proband and his parents图2 先证者及其父母SASH1 c.1574C>G变异位点的Sanger测序
2.3 预测T525R-SASH1 变异位点有害性和保守性
PolyPhen-2 软件预测,当SASH1基因第525 位苏氨酸(Thr)突变为精氨酸(Arg)时,Hum Div、Hum Var 数据集预测该突变对蛋白质结构可能有害(Probably damaging),预测分值分别为1 和0.99,预测结果提示该变异对蛋白质结构和功能造成影响的概率较大,见图3。MEGA7 软件对SASH1基因525位Thr 进行保守位点分析,结果显示Thr525 在不同物种间高度保守,见图4。
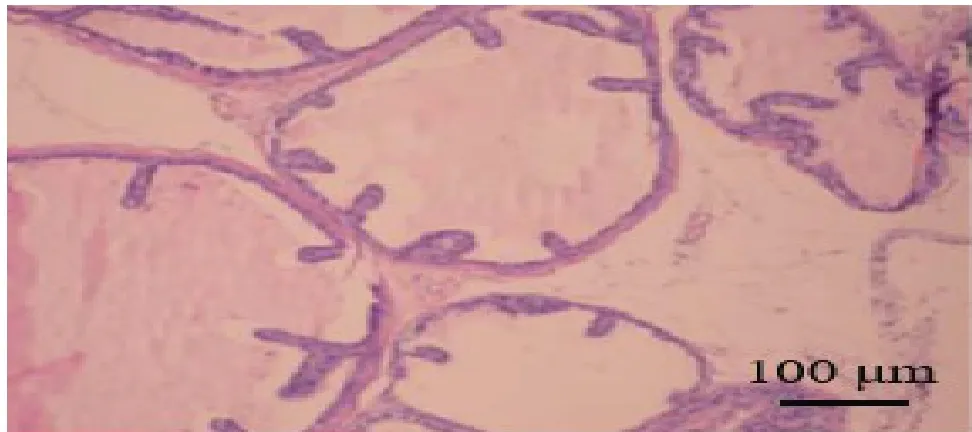
Fig. 3 PolyPhen-2 predicted the effect of T525R-SASH1 mutation on the structure and function of the protein图3 PolyPhen-2预测T525R-SASH1变异位点对该蛋白结构和功能的影响
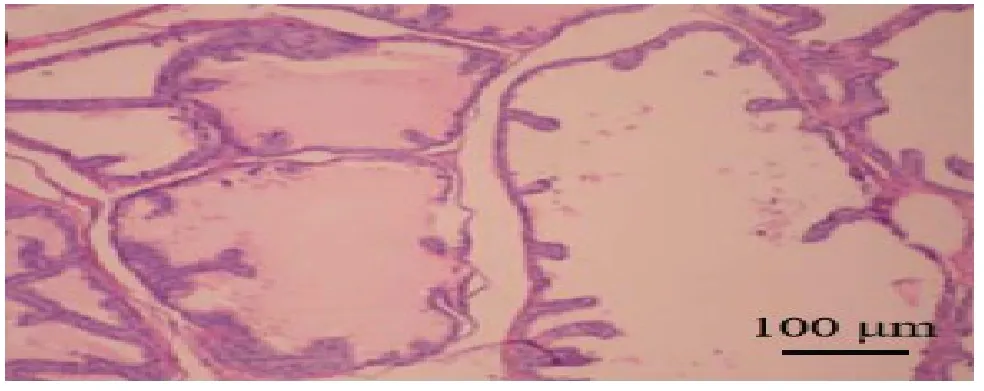
Fig. 4 MEGA7 predicted that the conserved nature of Thr525 locus in different species图4 MEGA7预测Thr525位点在不同物种之间的保守性
2.4 激光共聚焦显微镜明确SASH1 在PIG1 细胞的定位
通过细胞免疫荧光染色发现,正常人皮肤黑色素细胞PIG1表达SASH1蛋白,激光共聚焦显微镜观察到SASH1定位于PIG1细胞质中,见图5。
Fig.5 Confocal microscopy showed the localization of SASH1 in PIG1 cells(Immunofluorescence staining×200)图5 共聚焦显微镜显示SASH1在PIG1细胞中的定位(免疫荧光染色×200)
2.5 T525R-SASH1上调B16细胞MITF和Tyrosinase表达
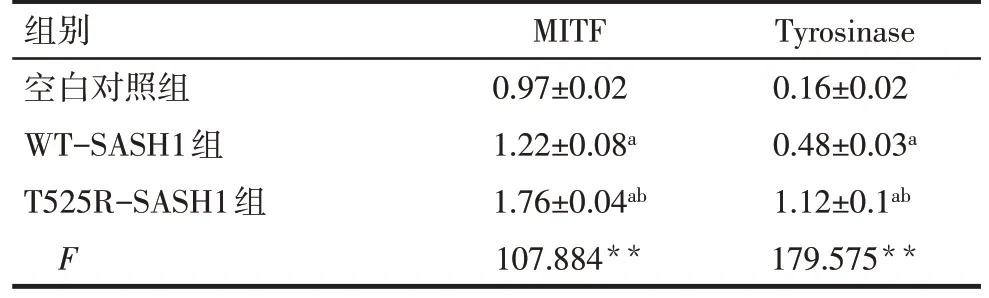
与空白组相比,过表达GFP-WT-SASH1和GFP-T525R-SASH1的 B16 细 胞 中 MITF 和Tyrosinase 的蛋白表达量增加(P<0.05),与过表达GFP-WT-SASH1的B16 细胞相比,过表达GFPT525R-SASH1细胞中MITF和Tyrosinase表达量增加(P<0.05),见图6、表1。
Tab. 1 Comparison of expression levels of MITF and Tyrosinase proteins between the three groups表1 3组MITF、Tyrosinase蛋白表达水平比较(n=3,±s)
Tab. 1 Comparison of expression levels of MITF and Tyrosinase proteins between the three groups表1 3组MITF、Tyrosinase蛋白表达水平比较(n=3,±s)
**P<0.01;a与空白对照组比较,b与WT-SASH1组比较,P<0.05。
组别空白对照组WT-SASH1组T525R-SASH1组F MITF 0.97±0.02 1.22±0.08a 1.76±0.04ab 107.884**Tyrosinase 0.16±0.02 0.48±0.03a 1.12±0.1ab 179.575**
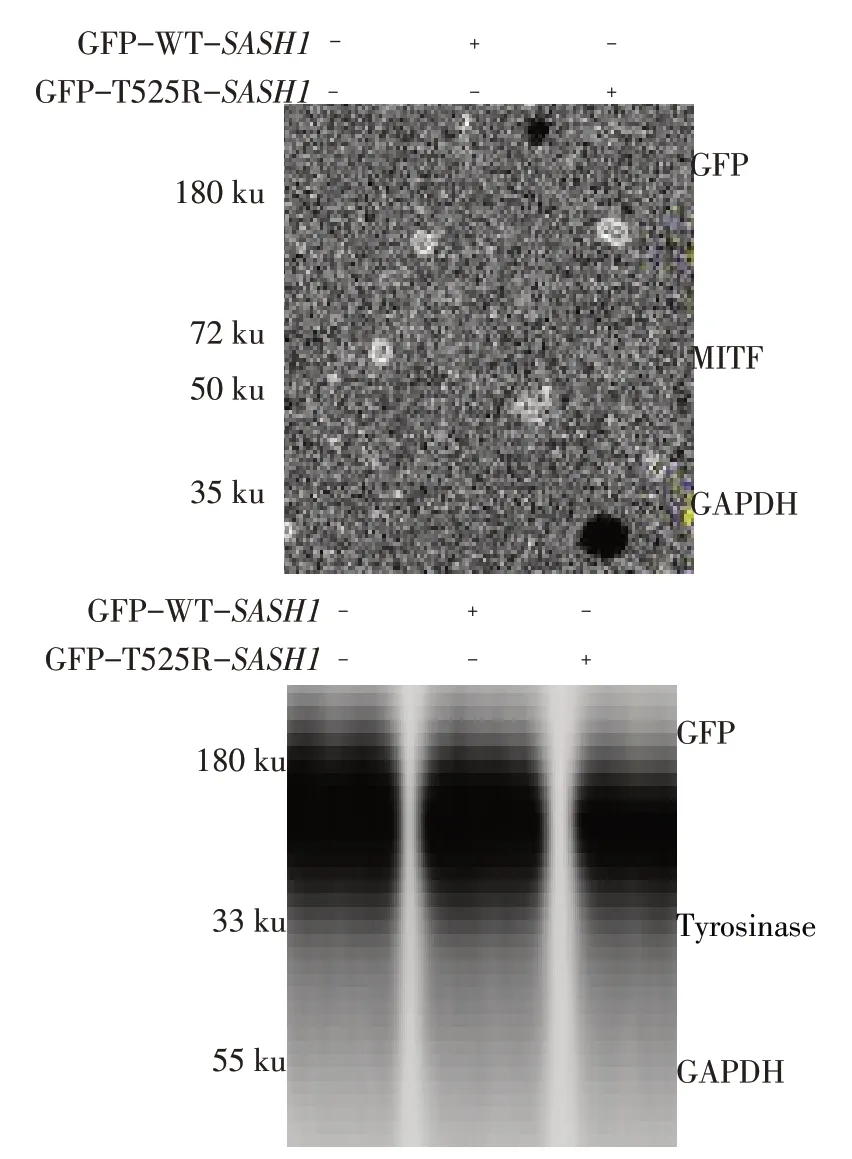
Fig.6 T525R-SASH1 overexpression increased MITF and Tyrosinase expression in B16 cells图6 T525R-SASH1上调B16细胞MITF和Tyrosinase蛋白表达
2.6 T525R-SASH1促进黑色素瘤细胞产生黑色素
Western blot 验证对照腺病毒组(NC ADV)、野生型SASH1腺病毒组(HA-WT-SASH1-ADV)、突变型T525R-SASH1腺病毒组(HA-MT-SASH1-ADV)成功过表达于B16 细胞(图7A)和SK-MEL-1 细胞(图7B)。提取成功过表达腺病毒的各组细胞的黑色素,与对照腺病毒组相比,过表达野生型SASH1腺病毒组和突变型SASH1腺病毒组的B16细胞产生的黑色素增加(P<0.05);过表达突变型SASH1腺病毒组的B16 细胞相比较野生型SASH1腺病毒组的B16细胞中产生的黑色素更多(P<0.05,图7C)。重复实验在SK-MEL-1细胞中得到一致的结果(图7D)。
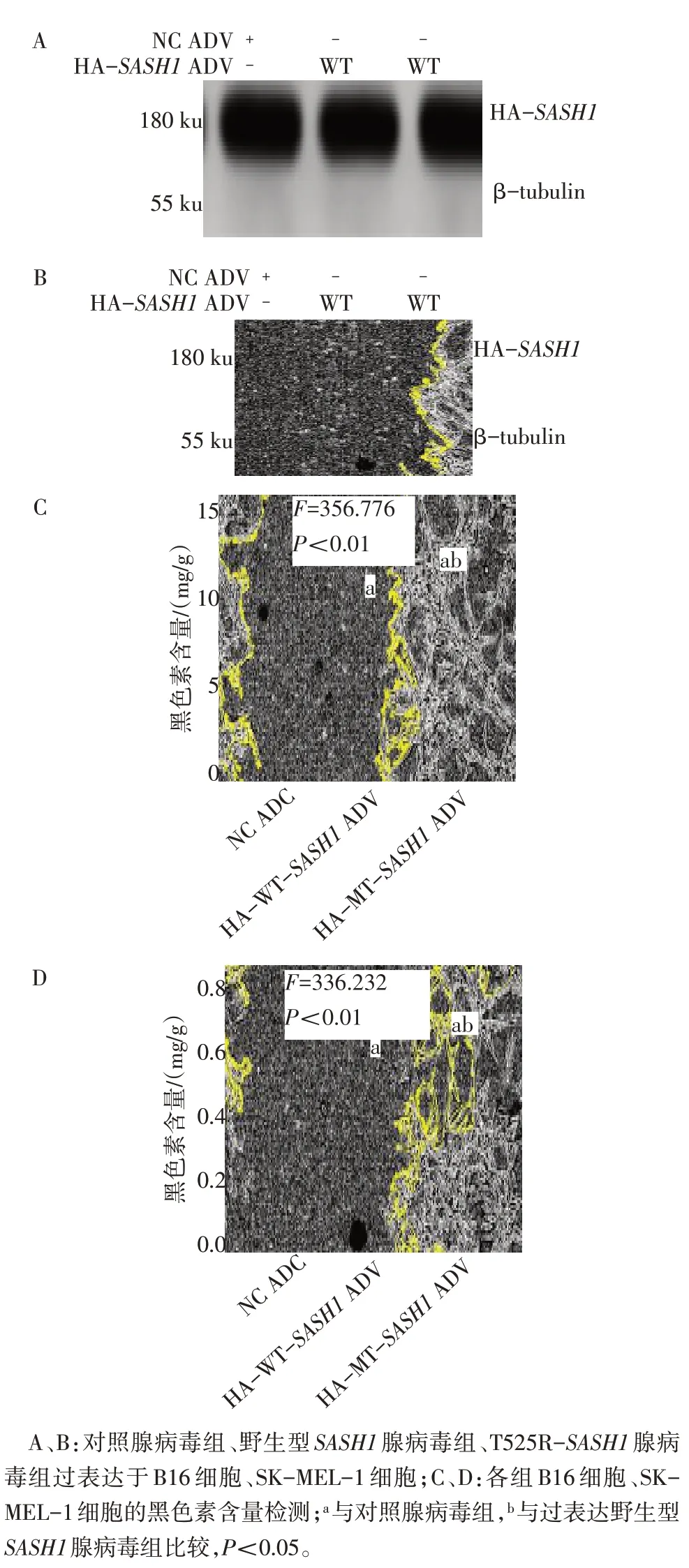
Fig.7 T525R-SASH1 over expression increased melanin synthesis in melanoma cells图7 过表达T525R-SASH1促进黑色素瘤细胞产生黑色素
3 讨论
3.1 SASH1 基因变异在DUH 发生中扮演着重要的角色
SASH1基因是2003 年德国学者Zeller 等[2]研究人类染色体6q23~25 的杂合性缺失时发现的,该基因位于染色体6q24.3,包含22个外显子和21个内含子,共279 746个碱基对,编码了1 230个氨基酸的蛋白,包含1 个SLY 结构域、1 个SH3 结构域和2 个SAM结构域。2003年Xing等[3]对2个汉族常染色体显性DUH 家系进行全基因组连锁定位,结果2 个家系的致病基因定位于染色体6q24.2~q25.2 区域,但未找到致病基因。2013年Zhou等[4]在研究2个中国的和1个美国的DUH家系时,首次在染色体6q24.2~6q25.2 区域发现SASH1基因SLY 结构域的3 个错义突变c. 2126T>G(p. Tyr551Asp),c. 2019T>C(p.Leu515Pro),c.2000G>A(p.Glu509Lys)与DUH 的表型发生相关。并先后证实这3个杂合性变异能够通过调节Gαs-SASH1-IQGAP1-E-cadherin[4]、p53-POMC-SASH1[5]和SASH1-MAP2K2-CREB-ERK[6]信号通路来调节患者皮肤色素过度沉着表型的发生。SASH1 的变异位点c.1553A >C p.Gln518Pro[7]、c. 1651T > C p. Tyr551His[8]、c. 1529G > A p.Ser510Asn[9]、c.1553A >C p.Gln518Pro[10]、c.1547G >A p.Ser516Asn[11]等被报道与DUH 发生相关,迄今为止文献共报道了12 个SASH1基因变异与DUH 发病相关。
3.2 SASH1 c.1574C>G(p.T525R)变异导致DUH 临床表型发生
全外显子组测序和Sanger测序检测发现先证者父母该变异位点均为野生型,先证者携带SASH1基因c.1574C>G(p.T525R)杂合变异是一个新发的变异位点。既往日本报道1例患者携带与本研究先证者相同的SASH1基因c.1574C>G(p.T525R)变异位点,但确诊为泛发性雀斑样痣[12],与本例患者的临床诊断和临床表现有较大差异,可见同一基因的同一变异位点可能会引起两种不同的疾病出现。
3.3 SASH1 c.1574C>G(p.T525R)杂合变异可促进黑色素合成增加
研究发现,以往报道与DUH相关的12个SASH1基因的变异位点中,有9 个变异位点均位于SASH1基因的SLY 结构域,该结构域高度保守,被视为DUH 的热点突变区域。本研究所报道SASH1c.1574C>G(p.T525R)变异位点正是位于高度保守的SLY结构域。PolyPhen-2软件预测SASH1基因第525位苏氨基酸突变为精氨酸后对蛋白质结构和功能造成影响的概率极高,野生型SASH1基因第525位苏氨酸在不同物种之间的保守性也高,所以推测SASH1c.1574C>G(p.T525R)变异可能导致了SLY结构域发生改变,从而影响SASH1蛋白的正常功能。本研究进一步通过体外功能验证该变异位点的致病性。黑色素合成的过程极其复杂,主要涉及酪氨酸酶基因家族中的3 种酶,即酪氨酸酶(Tyrosinase,TYR)、酪氨酸酶相关蛋白(Tyrosinase-related protein,TRP)1、TRP-2,其中TYR是黑色素合成的过程中的关键酶[13]。MITF 是一种碱性螺旋-环-螺旋亮氨酸拉链转录因子,可以同启动子区域的M-box基序结合之后调节TYR、TRP-1、TRP-2 的表达,从而调控黑色素合成[14]。人的皮肤颜色依赖于黑色素的合成,黑色素过量累积会导致色素沉着[15]。本研究利用B16 和SK-MEL-1 细胞构建过表达T525RSASH1的细胞系,发现T525R-SASH1可以上调在黑色素合成过程中最关键的TYR 以及MITF 的蛋白表达量,并且T525R-SASH1可以促进黑色素瘤细胞合成黑色素增加,体外功能实验结果推测T525RSASH1变异可能是先证者出现色素沉着斑的原因。
综上所述,本研究为SASH1基因变异相关联的DUH 患者出现色素沉着斑的临床表型提供了理论参考,并且丰富了与DUH相关的SASH1基因突变位点。体外功能实验进一步说明SASH1基因变异与黑色素合成有关,为DUH的基因诊断和治疗提供了有益信息。但SASH1c.1574C>G(p.T525R)变异位点如何调控黑色素生成的具体机制尚未阐述,因此课题组下一步将会扩大该患者家系,从信号通路和基因调控等方面进一步研究SASH1基因变异对黑色素生成的调节机制。



