冯 延,郭 嘉,许瑞勤,马思续,魏凤灵,张留君,杨国宇,夏平安,张改平(河南农业大学牧医工程学院,郑州 450002)
猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus, PRRSV)基因组全长约15.3 kb,为不分节段的单股正链RNA。目前在编码区中至少已经有10个开放阅读框(open reading frame, ORF)被证实,分别为ORF1a、ORF1b、ORF2a、ORF2b、ORF3、ORF4、ORF5、ORF6、ORF7和ORF5a,并且在部分开放阅读框之间存在一定的相互重叠[1]。ORF1占全基因全长的80%,可分为ORF1a和ORF1b,主要负责编码与病毒复制相关的复制酶等非结构蛋白[2]。ORF2~7位于病毒基因组3′端,负责编码病毒的结构蛋白,其中ORF5、ORF6、ORF7编码的囊膜糖蛋白(GP5)、基质蛋白(M)及核衣壳蛋白(N)为病毒的主要结构蛋白,是病毒构架的主要组成部分;ORF2a、ORF2b、ORF3、ORF4编码的囊膜有关的糖蛋白GP2a、GP2b、GP3、GP4为PRRSV的次要结构蛋白,而对于ORF5编码的GP5a蛋白的功能作用目前尚不完全清楚[1,3-4]。
目前PRRSV的GP2、GP3、GP4、GP5和M蛋白均被鉴定含有中和表位,其中GP5被认为是诱导中和抗体及交叉免疫保护的主要靶蛋白,一直以来都是PRRSV基因工程疫苗研究的热点基因,研究发现GP5至少含有5个B细胞表位和2个T细胞表位,可诱导中和抗体的产生,并且抗体的中和作用与血清中针对GP5的抗体滴度相关[5-6]。GP5蛋白的胞外区同时存在一个诱骗表位A(decay epitope, aa 27—30)和一个中和表位B(aa 37—45),A表位作为优势表位可以阻碍B表位的暴露并阻止B表位抗体的产生[7-8],其机制尚有待研究。然而诸多研究表明,以单独的GP5蛋白构建的基因工程亚单位疫苗,以及GP5/M构建的亚单位疫苗,虽然能够提供一定的免疫保护,但这种免疫保护只发生在同源毒株之间[6,9-10],并且保护能力有限,与弱毒疫苗株相比其诱导的免疫保护依然较弱。
猪是PRRSV的唯一天然宿主。在体外培养的众多细胞系中,PRRSV只在非洲绿猴肾细胞系MA104及其衍生Marc-145中生长。病毒在侵入宿主细胞时是通过细胞膜表面的受体分子与病毒表面的蛋白相互作用介导的。然而在有些情况下,抗体不但不能阻止病毒的感染,反而协助病毒侵入靶细胞,增强病毒的感染力和复制能力,这一现象就被称作抗体依赖性增强作用(antibody-dependent enhancement,ADE)。PRRSV就存在ADE现象[11],机体内抗PRRSV的抗体可以通过Fc受体结合到巨噬细胞表面,从而形成免疫复合物,使Fc受体阳性细胞病毒的感染作用增强,因此非中和抗体反而对PRRSV的感染起到促进作用[12]。PRRSV自然感染后机体针对GP5中和表位的抗体产生往往较迟且水平低下,不能及时有效地清除病毒,这些非中和抗体很可能会进一步介导PRRSV的ADE作用,造成持续性感染,这一特性也给疫苗的研究和使用造成了巨大困难。
目前,PRRSV GP5在诱导机体免疫保护中的作用是基于Marc-145细胞中和试验的研究结果,并不能真实反映出其在临床感染中的实际保护效果。因此,本研究旨在通过大肠杆菌和杆状病毒/昆虫表达系统分别表达PRRSV去掉跨膜区和信号肽的截短GP5蛋白以及通过杆状病毒/昆虫表达系统表达全长的GP5蛋白,进而制备抗血清并检测三种抗重组蛋白抗血清在PAM细胞上中和同源及异源PRRSV毒株的活性并分析其差异,从不同表达系统探讨GP5蛋白抗体的中和活性,以期为PRRSV亚单位疫苗的研究提供参考依据。
1 材料与方法
1.1 细胞、菌株、血清与质粒载体
大肠杆菌DH5α、BL21(DE3)、Bac-to-Bac杆状病毒表达系统供体质粒pFastBac-HTA、PRRSV VR2332株和HeN-3株(MegAlign分析两毒株GP5基因同源性为89.1%)、猪抗PRRSV VR2332阳性血清(PRRSV VR2332全病毒免疫健康猪制备保存,ELISA效价为1∶12 800)、鼠抗PRRSV VR2332阳性抗血清(PRRSV VR2332全病毒免疫小鼠制备保存,ELISA效价为1∶12 800)、Marc-145细胞、Sf9细胞均由河南农业大学兽医微生物实验室保存。感受态DH10Bac购自北京索莱宝科技有限公司(中国北京)。
1.2 主要试剂
2×TaqMix、2×Pfu Master Mix购自北京康为世纪生物科技有限公司(中国北京);限制性内切酶BamHⅠ和XhoⅠ、T4 DNA连接酶购自TaKaRa(中国大连),BAC质粒提取试剂盒购自Omega(美国);抗6×His单抗购自北京中杉金桥生物技术有限公司(中国北京);细胞培养基Sf-900ⅡSFM、转染试剂CellfectinⅡ Reagent购自Invitrogen(美国)。
1.3 PRRSV GP5蛋白的截短原核表达
利用Marc-145细胞增殖PRRSV VR2332 毒株(GenBank登录号:U87392.3),以其ORF5基因序列为模板,应用生物学软件Primer Premier 5.0设计一对扩增包含PRRSV ORF5全基因片段的特异引物(表1),通过RT-PCR方法扩增出完整的ORF5基因,连接至pMD18-T载体构建重组质粒pMD18T-ORF5并进行测序。根据ORF5基因的测序结果,使用分子生物学信息网站www.cbs.dtu.dk中的TMHMM工具预测GP5蛋白的跨膜区分别位于第7—29、66—88和107—125位氨基酸。同时以SignalP 4.1 Server预测其信号肽位于1—31 aa。通过重叠延伸PCR技术对GP5蛋白的信号肽和跨膜区进行缺失处理,表达第32—65 aa的近N-端序列和第126—200 aa的近C-端序列(以下分别简称ORF5N和ORF5C),设计两对引物(表1),在ORF5NR与ORF5CF之间包含25 bp的反向互补序列(以下划线标注),并在两个多肽之间引入了3个串联的柔性氨基酸(Gly-Ser-Ser)序列以消除因多肽的串联对其构象造成的影响(以黑体标注),扩增出缺失跨膜区(aa 66—125)和信号肽(aa 1—31)序列的ORF5基因片段并连接至原核表达载体pET-32a,构建原核表达载体pET-ORF5NC。将重组质粒转化至大肠杆菌BL21中表达蛋白。
1.4 PRRSV GP5蛋白的全长和截短蛋白的真核表达
通过杆状病毒/昆虫表达系统分别表达全长的重组GP5蛋白和缺失跨膜区、信号肽的重组GP5蛋白。根据1.3所获得的ORF5基因片段,再次设计两对引物(表1),其中一对引物以质粒pD18T-ORF5为模板,扩增出完整的ORF5基因片段(ORF5-1);另一对引物以质粒pET-ORF5NC为模板,扩增出缺失跨膜区、信号肽的ORF5基因片段(ORF5NC),分别连接至pFastBac-HTA供体质粒,将获得的两种重组供体质粒分别转化至含有杆状病毒穿梭质粒的DH10-Bac细胞,经三重抗性和蓝白斑筛选,获得两种重组穿梭质粒Bacmid-ORF5和Bacmid-ORF5NC。在脂质体转染试剂Cellfectin Ⅱ Reagent的介导下,将重组Bacmid穿梭载体转染Sf9细胞,获得重组杆状病毒AcMNPV-ORF5和AcMNPV-ORF5NC;将两种重组杆状病毒分别感染Sf9细胞以期获得蛋白的表达。
1.5 三种重组蛋白抗血清以及PAM细胞的制备
将三种重组蛋白作为免疫原,分别免疫昆明小鼠制备三种抗血清。每只小鼠抗原用量为75 μg(200 μL),初次免疫时将纯化的重组蛋白与弗氏完全佐剂按1∶1.2的体积分别乳化均匀,首免2周后使用弗氏不完全佐剂与重组蛋白乳化均匀,采用背部皮下多点注射小鼠,以后每隔一周免疫一次,共免疫4次。以各重组蛋白和PRRSV全病毒作为ELISA包被抗原,检测抗血清的效价。
从PRRSV核酸与抗体双阴性的4周龄健康仔猪中无菌摘取肺,制备肺泡巨噬细胞悬液,500 g离心20 min,弃去上清,细胞以含有1 000 IU·L-1青、链霉素的PBS液轻柔重悬洗涤2次,最后将PAM细胞重悬于含10%胎牛血清的RPMI1640培养液中。
表1引物序列及相关信息
Table1Primersequencesandrelevantinformation

扩增片段名称Amplifiedfragmentname引物序列(5′→3′)Primersequences酶切位点Restrictionenzyme扩增片段长度/bpAmplifiedfragmentlengthORF5ORF5F:ATGTTGGAGAAATGCTTGACCG603ORF5R:CTAAGGACGACCCCATTGTTCORF5NORF5NF:CGCGGATCCAGCAACGACAGCAGCTCCCATBamHⅠ127ORF5NR:GCAAACCTGGAAGAGCCCTCCACTGCCCAATCORF5CORF5CF:CAGTGGAGGGCTCTTCCAGGTTTGCAAAGAAT248ORF5CR:CCGCTCGAGAGGACGACCCCATTGTTCCGXhoⅠORF5⁃1ORF5F:CGCGGATCCGATGTTGGAGAAATGCTTGACCGBamHⅠ618ORF5R:CCGCTCGAGACCTAAGGACGACCCCATTGTTCXhoⅠORF5NCORF5NF:CGCGGATCCGAGCAACGACAGCAGCTCCCATBamHⅠ353ORF5CR:CCGCTCGAGACAGGACGACCCCATTGTTCCGXhoⅠ
下划线表示反向互补序列,黑体部分为柔性氨基酸
Underline represents reverse complementary sequence, the blackbody part represents the flexible amino acid
调整细胞浓度至106·mL-1。向24孔细胞培养板中每孔加入0.5 mL的细胞悬液,并轻摇细胞培养板使细胞均匀分布,于37 ℃、5% CO2的细胞培养箱中培养6 h使细胞贴壁,而后弃去未贴壁的细胞,以灭菌的PBS轻柔洗涤细胞一次,每孔加入0.5 mL的含10%胎牛血清的RPMI1640培养液,置于37 ℃、5% CO2的细胞培养箱中培养。
1.6 抗GP5重组蛋白抗体在PAM细胞上的中和活性分析
将达到效价的抗重组蛋白血清和阴性血清灭活处理后,均做2、4、8、16、32倍的系列稀释,分别与等体积的含200个TCID50的同源毒株PRRSV VR2332和异源毒株PRRSV HeN-3作用1 h后,接种于单层的PAM细胞,同时设立鼠PRRSV阳性血清对照、接毒细胞对照、正常细胞对照和血清毒性对照。48 h后收获样品,以段二珍[13]建立的PRRSV实时定量PCR方法检测各孔中PRRSV拷贝数;采用双因素方差分析法(Two-way ANOVA)对各组数据进行分析比较,以王冬梅[14]建立的中和抗体检测方法确定中和活性。
2 结 果
2.1 PRRSV 截短GP5蛋白的原核表达
将成功构建的原核表达载体pET-ORF5NC重组质粒转化至表达菌BL21中,以最佳的IPTG诱导浓度和最佳的诱导表达时间成功表达出大小约31 ku的截短重组GP5蛋白(N-端融合有大小为17.6 ku的6×His蛋白标签),与预期蛋白大小相符。以尿素法纯化包涵体蛋白并复性,分别以鼠抗His单克隆抗体和PRRSV阳性血清为一抗对重组蛋白进行Western blotting检测,结果显示,重组蛋白可以被鼠抗His单克隆抗体和猪抗PRRSV阳性血清(图1: 7、8泳道)所识别,表明重组蛋白具有良好的反应原性。
2.2 PRRSV 全长和截短GP5蛋白的真核表达
两种重组蛋白在Sf9细胞中成功获得了不溶性表达。以SDS-PAGE和Western blotting对两种重组杆状病毒所表达的蛋白进行鉴定,结果表明,Sf9细胞中分别表达了大小约为22 ku的全长GP5重组蛋白和大小为17 ku的截短GP5重组蛋白,与预期蛋白大小相符,正常细胞和野生型病毒感染细胞对照中均未检测到特异性条带。大量表达的两种重组蛋白经Ni+柱亲和层析方法进行纯化,并通过SDS-PAGE以及ImageJ软件进行灰度分析,其中AcMNPV-ORF5表达纯化产物为90.3%,AcMNPV-ORF5表达纯化产物为92.6%。同时,分别以抗His单克隆抗体和猪抗PRRSV阳性血清对重组蛋白进行Western blotting分析(图1: 1~6泳道)。
2.3 抗不同类型PRRSV GP5重组蛋白抗体的中和活性分析
2.3.1 三种重组蛋白抗血清的ELISA效价检测 以方阵滴定法对三种重组蛋白的最佳抗原包被浓度进行摸索,发现原核截短的GP5蛋白最佳包被浓度为5 μg·mL-1,真核全长和真核截短的GP5蛋白的最佳包被浓度均为2.5 μg·mL-1。以最佳的重组蛋白抗原包被浓度包被ELISA平板,检测三种重组蛋白抗血清的效价,以待检测孔OD450 nm≥0.3且P/N值≥2.1作为判定标准,测得抗原核截短GP5蛋白、抗真核全长GP5蛋白和抗真核截短GP5蛋白抗血清的ELISA效价分别为1∶25 600、1∶12 800和1∶12 800。当以PRRSV全病毒作为ELISA包被抗原检测抗血清时,测得抗原核截短GP5蛋白、抗真核全长GP5蛋白和抗真核截短GP5蛋白抗血清的ELISA效价分别为1∶200、1∶800和1∶1 600。结果表明,三种重组蛋白抗血清均具有良好的反应原性。

M. 蛋白质相对分子量标准;1. 以6×His单抗检测重组蛋白AcMNPV-ORF5;2. 以猪PRRSV VR2332阳性血清检测重组蛋白AcMNPV-ORF5;3. 正常Sf9细胞总蛋白;4. 以6×His单抗检测重组蛋白AcMNPV-ORF5NC;5. 以猪PRRSV阳性血清检测重组蛋白AcMNPV-ORF5NC;6. 经野生型杆状病毒感染后的Sf9细胞总蛋白;7. 以6×His单抗检测纯化的原核表达GP5重组蛋白;8. 以猪抗PRRSV VR2332阳性血清检测纯化的原核表达GP5重组蛋白M. Protein marker; 1. Recombinant protein of AcMNPV-ORF5 with anti-6×His mAb as the first antibody; 2. Recombinant protein of AcMNPV-ORF5 with PRRSV VR2332 porcine serum as the first antibody; 3. Total cellular proteins of Sf9; 4. Recombinant protein of AcMNPV-ORF5NC with anti-6×His mAb as the first antibody; 5. Recombinant protein of AcMNPV-ORF5NC with PRRSV porcine serum as the first antibody; 6. Total cellular proteins of Sf9 infected with AcMNPV; 7. Recombinant GP5 protein through prokaryotic expression system with anti-6×His mAb as the first antibody; 8. Recombinant GP5 protein through prokaryotic expression system with PRRSV VR2332 porcine serum as the first antibody
图1 以6×His单抗和猪抗PRRSV VR2332阳性血清检测GP5重组蛋白的Western blotting结果Fig.1 Western blotting analysis of recombinant proteins by anti-6×His mAb and PRRSV VR2332-positive serum
2.3.2 抗三种GP5重组蛋白抗体对同源株PRRSV VR2332的中和活性 结果表明,三种抗血清均不能中和同源株PRRSV VR2332对PAM细胞的感染。虽然真核截短GP5抗血清在1∶2和1∶4倍稀释时对同源病毒复制表现出略微的抑制性,但是相比阴性血清并没有显着差异;原核截短GP5蛋白抗血清在1∶2 倍稀释、真核全长GP5蛋白抗血清在1∶2倍稀释、真核截短GP5蛋白抗血清在1∶8、1∶16、1∶32倍稀释时显着增强了同源病毒的感染能力(图2a~c)。对照组的鼠抗PRRSV VR2332阳性抗血清可以中和同源株PRRSV VR2332的感染,中和效价为1∶8(图2d)。
2.3.3 抗三种GP5重组蛋白抗体对异源株PRRSV HeN-3的中和活性 结果显示,三种抗血清均不能中和异源株PRRSV HeN-3对PAM细胞的感染。虽然真核截短GP5蛋白抗血清在1∶2倍稀释、真核全长GP5蛋白抗血清在所有系列稀释时对异源病毒复制都表现出略微的抑制性,但是相比阴性血清并没有显着差异(图3a~c)。原核截短GP5蛋白抗血清反而显着增强了异源毒株的感染能力,真核截短GP5蛋白在1∶4倍稀释时也显着增强了病毒的感染。对照组的PRRSV阳性抗血清不可以中和异源株PRRSV HeN-3的感染,反而使病毒的感染能力增强(图3d)。
3 讨 论
PRRSV GP5蛋白长期以来被认为是诱导中和抗体产生的主要蛋白[15]。国内外的相关研究均报道了中和抗体在抵抗PRRSV感染中的重要作用[16-17],制备诱导高滴度中和抗体的疫苗在PRRSV的预防上具有重要意义。以杆状病毒系统表达GP5蛋白用于PRRSV亚单位疫苗的研究也有一定的报道[18]。
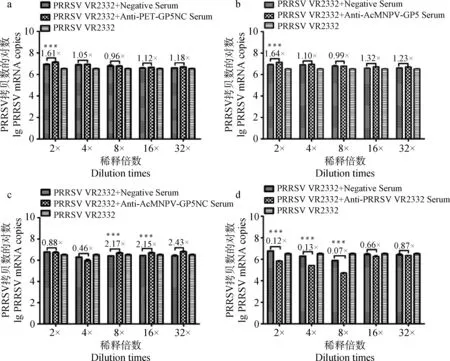
a. 抗原核截短GP5蛋白抗血清中和同源株PRRSV VR2332活性的检测;b. 抗真核全长GP5蛋白抗血清中和同源株PRRSV VR2332活性的检测;c. 抗真核截短GP5蛋白抗血清中和同源株PRRSV VR2332活性的检测;d. 鼠源抗PRRSV VR2332阳性血清中和同源株PRRSV VR2332活性的检测;***.P<0.001a. Neutralizing activity of mouse anti-pET-GP5NC serum against PRRSV VR2332; b. Neutralization activity of mouse anti-AcMNPV-GP5 serum against PRRSV VR2332; c. Neutralization activity of mouse anti-AcMNPV-GP5NC serum against PRRSV VR2332; d. Neutralization activity of mouse anti-PRRSV VR2332 serum against PRRSV VR2332;***.P<0.001
图2 三种GP5重组蛋白抗血清与同源株PRRSV VR2332的中和活性比较Fig.2 Neutralizing activity of mouse sera to the three recombinant proteins against PRRSV VR2332
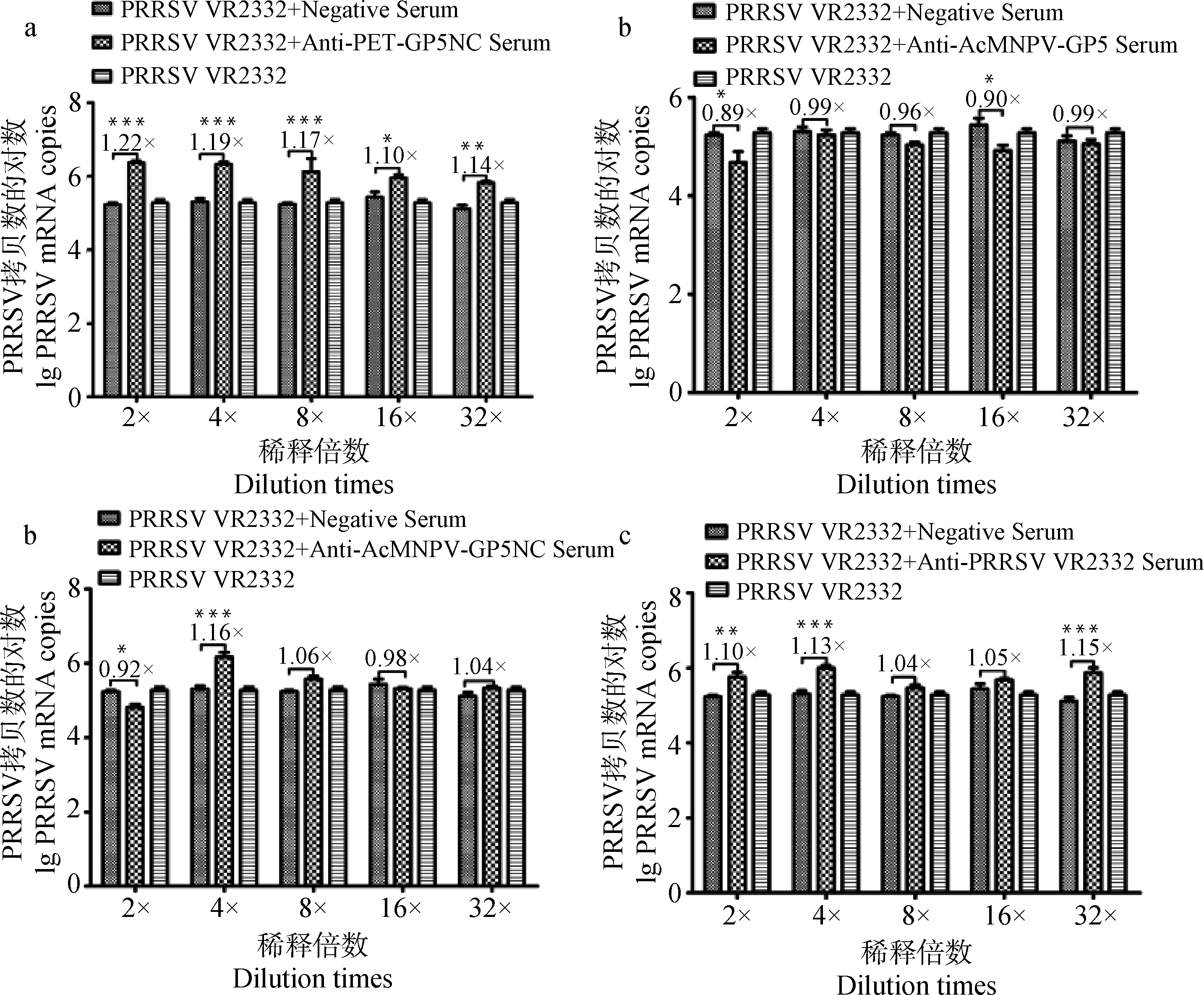
a.抗原核截短GP5蛋白抗血清中和异源株PRRSV HeN-3活性的检测;b.抗真核全长GP5蛋白抗血清中和异源株PRRSV HeN-3活性的检测;c.抗真核截短GP5蛋白抗血清中和异源株PRRSV HeN-3活性的检测;d.鼠源抗PRRSV VR2 332阳性血清中和异源株PRRSV HeN-3活性的检测;*.P<0.05;**.P<0.01;***.P<0.001a. Neutralizing activity of mouse anti-pET-GP5NC serum against PRRSV HeN-3; b. Neutralizing activity of mouse anti-AcMNPV-GP5 serum against PRRSV HeN-3; c. Neutralizing activity of mouse anti-AcMNPV-GP5NC serum against PRRSV HeN-3; d. Neutralizing activity of mouse anti-PRRSV VR2332 serum against PRRSV HeN-3;*.P<0.05;**.P<0.01;***.P<0.001
图3 三种GP5重组蛋白抗血清与异源株PRRSV HeN-3的中和活性比较Fig.3 Neutralizing activity of mouse sera to the three recombinant proteins against PRRSV HeN-3
Xu等[19]以杆状病毒表面展示系统将PRRSV GP5蛋白和猪圆环病毒2型(porcine circovirus type 2,PCV2)的衣壳蛋白共展示在杆状病毒囊膜表面,将重组病毒免疫猪,免疫血清中可检测到高水平的抗GP5蛋白的ELISA抗体,但具有较低水平的中和活性;早期的研究发现,在一定量补体存在的情况下,感染PRRSV后康复期的猪血清可部分中和PRRSV对Marc-145细胞的感染[20]。但以上研究均是基于Marc-145细胞的中和试验结果。由于临床上PRRSV只感染PAM细胞,而PAM细胞同时含有Marc-145细胞所不具有的Fc受体和唾液酸黏附素受体(Sn/CD169),可以介导PRRSV 的ADE效应,所以,基于Marc-145细胞的中和试验结果并不能真实反映其抗体的中和作用,因此有必要基于PAM细胞的中和试验来研究抗GP5蛋白抗体的中和活性。本试验通过杆状病毒/昆虫表达系统分别表达了全长和缺失信号肽、跨膜区的截短GP5蛋白,同时用大肠杆菌表达系统表达了截短的GP5蛋白,通过免疫小鼠获得抗血清,并将三种抗GP5蛋白血清及抗PRRSV全病毒血清在PAM细胞上进行了同源和异源毒株的中和试验,结果显示,虽然三种GP5重组蛋白的抗血清ELISA效价较高,但均没有检测到其中和活性,而抗全病毒的阳性抗血清,如果按照100%抑制标准,对同源和异源毒株也没有中和活性,如果按照70%抑制标准,对同源毒株具有部分中和活性,效价为1∶8,对异源毒株的中和效价<1∶2,甚至还介导了ADE效应,增强了病毒感染,这与基于Marc-145细胞的中和试验研究结果并不完全一致,说明了基于Marc-145细胞的中和试验研究结果与实际情况存在明显差异。
最近的一些研究表明,GP5蛋白的主要中和表位可能更多是构象表位[6, 21-22],鉴于人工表达的GP5重组蛋白无法获得构象型中和抗原表位,其诱导的抗体无疑不具有有效的中和活性。本研究结果显示真核表达的抗全长和截短GP5蛋白血清均无中和活性,而PRRSV全病毒阳性抗血清对同源毒株的感染具有部分中和活性,提示PRRSV全病毒中主要诱导中和抗体产生的中和表位并不完全在GP5蛋白上,目前被鉴定存在中和表位的PRRSV蛋白还有GP2a、GP3、GP4和M蛋白[9, 23-25]。另外,GP5蛋白的信号肽中存在有高度变异的诱骗表位A,其作为优势表位可以阻碍中和表位B的暴露并阻止B表位抗体的产生[7-8],这可能也是影响真核表达全长GP5蛋白抗血清免疫原性的原因之一。本试验中三种GP5重组蛋白的部分浓度抗血清还可以增加病毒的复制水平,可能是GP5重组蛋白在免疫小鼠时产生的非中和抗体所占比重较大,促使了PRRSV的ADE效应产生,致使特异性的中和抗体无法发挥中和作用。本研究中的抗PRRSV全病毒阳性血清对异源毒株不具有中和活性,反而增强了病毒的感染,说明PRRSV诱导的抗体虽然能够提供一定的免疫保护,但这种免疫保护只发生在同源毒株之间,对异源毒株并没有效果,反而会介导ADE效应发生。
4 结 论
利用三种不同形式的猪繁殖与呼吸综合征病毒GP5重组蛋白抗血清和PRRSV全病毒高免血清在PAM细胞上进行了同源和异源毒株的中和试验,结果显示三种GP5重组蛋白抗血清没有中和活性,全病毒抗血清对同源毒株具有部分中和作用,而对异源毒株则没有中和活性,反而会介导ADE效应发生,研究表明PRRSV诱导产生中和抗体的中和表位并不完全在GP5蛋白上,而在GP5蛋白上的中和表位可能更多是构象表位,这将为以GP5为目标基因制备PRRSV亚单位疫苗的研究提供进一步参考。
参考文献(References):
[1] FIRTH A E, ZEVENHOVEN-DOBBE J C, WILLS N M, et al.Discovery of a small arterivirus gene that overlaps the GP5 coding sequence and is important for virus production[J].JGenVirol, 2011, 92(Pt 5):1097-1106.
[2] JOHNSON C R,GRIGGS T F,GNANANDARAJAH J,et al.Novel structural protein in porcine reproductive and respiratory syndrome virus encoded by an alternative ORF5 present in all arteriviruses[J].JGenVirol, 2011,92(Pt 5):1107-1116.
[3] MEULENBERG J J M,VAN NIEUWSTADT A P,VAN ESSEN-ZANDBERGEN A,et al.Posttranslational processing and identification of a neutralization domain of the GP4protein encoded by ORF4 of Lelystad virus[J].JVirol,1997,71(8):6061-6067.
[4] SNIJDER E J,VAN TOL H,PEDERSEN K W,et al.Identification of a novel structural protein of arteriviruses[J].JVirol,1999,73(8):6335-6345.
[5] GONIN P, PIRZADEH B, GAGNON C A,et al.Seroneutralization of porcine reproductive and respiratory syndrome virus correlates with antibody response to the GP5major envelope glycoprotein[J].JVetDiagnInvest, 1999, 11(1):20-26.
[6] PIRZADEH B,DEA S.Immune response in pigs vaccinated with plasmid DNA encoding ORF5 of porcine reproductive and respiratory syndrome virus[J].JGenVirol,1998,79(Pt 5):989-999.
[7] PLAGEMANN P G W.The primary GP5 neutralization epitope of North American isolates of porcine reproductive and respiratory syndrome virus[J].VetImmunolImmunopathol,2004,102(3):263-275.
[8] QIU H J,TIAN Z J,TONG G Z,et al.Protective immunity induced by a recombinant pseudorabies virus expressing the GP5 of porcine reproductive and respiratory syndrome virus in piglets[J].VetImmunolImmunopathol,2005,106(3-4):309-319.
[9] VANHEE M,VAN BREEDAM W,Costers S,et al.Characterization of antigenic regions in the porcine reproductive and respiratory syndrome virus by the use of peptide-specific serum antibodies[J].Vaccine,2011,29(29-30):4794-4804.
[10] ZHOU J X,XUE J D,YU T,et al.Immune responses in pigs induced by recombinant canine adenovirus 2 expressing the glycoprotein 5 of porcine reproductive and respiratory syndrome virus[J].VetResCommun,2010,34(4):371-380.
[11] YOON K J,WU L L,ZIMMERMAN J J,et al.Antibody-dependent enhancement (ADE) of porcine reproductive and respiratory syndrome virus (PRRSV) infection in pigs[J].ViralImmunol,1996,9(1):51-63.
[12] WILLS R W,DOSTER A R,GALEOTA J A,et al.Duration of infection and proportion of pigs persistently infected with porcine reproductive and respiratory syndrome virus[J].JClinMicrobiol,2003,41(1):58-62.
[13] 段二珍.PRRSV的感染差异性和抗体依赖性增强作用研究[D].郑州:河南农业大学,2011.
DUAN E Z.Studies on the difference of PRRSV infectious and PRRSV ADE[D].Zhengzhou:Henan Agricultural University,2011.(in Chinese)
[14] 王冬梅.不同免疫原对PRRSV诱导中和抗体的影响及SHP1在PRRSV感染中的作用[D].郑州:河南农业大学,2015.
WANG D M.The effect of different immunogens on PRRSV-induced neutralizing antibody and role of SHP1 in PRRSV infection[D].Zhengzhou:Henan Agricultural University,2015.(in Chinese)
[15] LUNNEY J K,FANG Y,LADINIG A,et al.Porcine reproductive and respiratory syndrome virus (PRRSV):Pathogenesis and interaction with the immune system[J].AnnuRevAnimBiosci,2016,4:129-154.
[16] LOPEZ O J,OLIVEIRA M F,GARCIA E A,et al.Protection against porcine reproductive and respiratory syndrome virus (PRRSV) infection through passive transfer of PRRSV-neutralizing antibodies is dose dependent[J].ClinVaccineImmunol,2007,14(3):269-275.
[17] 夏平安,崔保安,张红英,等.特异性中和抗体终止猪体内猪繁殖与呼吸综合征病毒(PRRSV)复制[J].中国兽医学报,2006,26(4):354-356.
XIA P A,CUI B A,ZHANG H Y,et al.Specific neutralizing antibodies terminate replication of porcine reproductive and respiratory syndrome virus (PRRSV) in PRRSV infected pigs[J].ChineseJournalofVeterinaryScience,2006,26(4):354-356.(in Chinese)
[18] URNIZA A,CLIMENT J,DURAN J P,et al.Baculovirus expression of proteins of porcine reproductive and respiratory syndrome virus strain Olot/91.Involvement of ORF3 and ORF5 proteins in protection[J].VirusGenes,1997,14(1):19-29.
[19] XU X G,WANG Z S,ZHANG Q,et al.Baculovirus as a PRRSV and PCV2 bivalent vaccine vector:baculovirus virions displaying simultaneously GP5 glycoprotein of PRRSV and capsid protein of PCV2[J].JVirolMethods,2012,179(2):359-366.
[20] YOON I J,JOO H S,GOYAL S M,et al.A modified serum neutralization test for the detection of antibody to porcine reproductive and respiratory syndrome virus in swine sera[J].JVetDiagnInvest,1994,6(3):289-292.
[21] LI J,MURTAUGH M P.Dissociation of porcine reproductive and respiratory syndrome virus neutralization from antibodies specific to major envelope protein surface epitopes[J].Virology,2012,433(2):367-376.
[22] FAN B C,LIU X,BAI J,et al.The amino acid residues at 102 and 104 in GP5 of porcine reproductive and respiratory syndrome virus regulate viral neutralization susceptibility to the porcine serum neutralizing antibody[J].VirusRes,2015,204:21-30.
[23] OSTROWSKI M,GALEOTA J A,JAR A M,et al. Identification of neutralizing and nonneutralizing epitopes in the porcine reproductive and respiratory syndrome virus GP5 ectodomain[J].JVirol,2002,76(9):4241-4250.
[24] YANG L,FREY M L,YOON K J,et al.Categorization of North American porcine reproductive and respiratory syndrome viruses:epitopic profiles of the N,M,GP5 and GP3 proteins and susceptibility to neutralization[J].ArchVirol,2000,145(8):1599-1619.
[25] TRIBLE B R,POPESCU L N,MONDAY N.A single amino acid deletion in the matrix protein of porcine reproductive and respiratory syndrome virus confers resistance to a polyclonal swine antibody with broadly neutralizing activity[J].JVirol,2015,89(12):6515-6520.



