罗 菊,毛嘉妮,夏银钊,杨震国
(西南大学动物科学技术学院 生物饲料与分子营养实验室,重庆 400715)
在机体消化过程中,肠黏膜选择性地吸收水和营养物质、阻止肠道细菌异位、发挥肠道免疫功能,以协助机体维持肠道稳态[1]。肠道是机体内环境与外部环境相互作用的门户,它不仅是重要的消化吸收场所也是机体重要的免疫防御屏障。功能上,肠道黏膜屏障可分为机械屏障、化学屏障、免疫屏障和生物屏障[2]。多种因素均可导致肠道屏障功能障碍,完整的肠道屏障对于机体健康的维持和疾病预防至关重要[3]。
现有研究表明,circRNAs在哺乳动物肠道中发挥重要调控作用,并通过多种方式参与调控肠道屏障的维持。Zeng等[4]检测了猪乳外泌体(porcine milk exosomes, PME)中长链非编码RNA(long non-coding RNAs, lncRNAs)和circRNAs的表达情况,结果显示PME中共鉴定出61个circRNAs,进一步探究发现circRNAs可能靶向许多与肠道屏障相关的miRNAs。Chung等[5]敲除小鼠肠道中circRNACdr1as后发现,小鼠肠道黏膜组织中潘氏细胞数量增加,此外,circRNACdr1as缺失后肠道的机械屏障功能增强。Li等[6]研究发现,hsa_circ_0001021在结肠炎患者中表达显着下调,hsa_circ_0001021可与miR-224-5p相互作用调节肠道上皮屏障功能。目前关于circRNAs的研究多集中在疾病防治、组织特异性、配子发生及早期胚胎发育等医学和生物学领域,对于其在哺乳动物肠道屏障中所发挥的调控机制研究还处于起步阶段,在家畜中的相关研究更是鲜有报道。本文主要综述了circRNAs在哺乳动物肠道机械屏障、肠道免疫屏障及肠道微生物屏障中的调控机制,以期为其在畜牧生产实践中能够加以运用提供理论依据。
1 circRNAs概况
1.1 circRNAs的发现
与线性RNA不同,circRNAs是一类由非编码RNA的5′和3′端共价连接形成的闭合circRNAs[7]。1976年,Sanger等[8]首次在植物感染的类病毒中发现了这些呈闭合环状的单链RNA分子。虽然在真核生物中也发现了circRNAs存在,但是由于当时技术的限制,circRNAs被认为是一种RNA转录剪切错误未能引起重视。近年来,随着高通量测序等技术的发展,许多circRNAs被发现与人类癌症、肿瘤等疾病有关,circRNAs成为了新的研究热点。
1.2 circRNAs的特点
据目前的研究来看,circRNAs具有以下特点:1)circRNAs广泛存在于真核生物细胞中[9];2)circ-RNAs具有高度保守性,多数具有高度保守序列,仅少数会产生进化改变[10];3)circRNAs大部分是ncRNAs,呈闭合环状;4)circRNAs不具有5′末端帽子和3′末端尾巴,不易被RNA核糖核酸外切酶R(ribonuclease R,RNase R)破坏,比线性RNA稳定性更好[11];5)大量circRNAs存在于细胞质中,少部分位于细胞核中[11];6)circRNAs大多数来源于外显子,少数来源于内含子[12];7)部分circRNAs具有miRNAs应答元件,与miRNAs相互作用,调控其靶基因的表达[13];8)大部分circRNAs能在转录或转录后水平发挥调控作用,少数只能在转录水平发挥作用[14]。
1.3 circRNAs的分类
按照circRNAs序列来源,可分为以下4类:1)源于外显子的circRNAs (exonic circRNAs,ecirc RNAs):仅由外显子组成,主要存在于细胞质中,含量最丰富[15-16];2)源于内含子的circRNAs(circular intronic RNAs,ciRNAs):由内含子组成,主要存在于细胞核中[17];3)由外显子和内含子共同构成的circRNAs(exon-intron circRNAs,EIcircRNAs):由外显子和内含子组成,主要在细胞核中表达[16,18];4)病毒circRNAs,由病毒RNA基因组、tRNAs、rRNAs和snRNAs等的环状化产生[19]。
1.4 circRNAs的产生机制
根据不同的环化方式,circRNAs的产生机制可分为以下3种类型:1)由套索驱动环化形成的circRNAs:该途径是在mRNA前体上游5′剪接供体位点与下游3′剪接受体位点以共价键结合,产生含有外显子和内含子的套索,再通过反向剪接切除内含子,产生circRNAs[20];2)由内含子配对驱动环化形成的circRNAs:在该途径中circRNAs外显子两侧的内含子存在反向互补序列,内含子在剪接位点反向互补配对,随后将互补配对的内含子反向剪接,由此形成不同种类型的circRNAs[13];3)RNA结合蛋白(RNA-binding protein,RBP)驱动环化形成的circRNAs,RBP通过与circRNAs外显子两端的内含子结合从而拉近了供体位点和受体位点的距离,促进circRNAs的产生[21]。
1.5 circRNAs的功能
研究发现,circRNAs主要有以下功能:1)充当miRNAs的海绵;miRNAs是一种长度约20~24个核苷酸的小RNA,可以通过与mRNA的5′和3′端非编码区(untranslate dregions,UTRs)结合,使目标mRNA降解,广泛参与转录后基因表达调控作用[22-23]。circRNAs上存在结合位点可与特定miRNAs结合,减少miRNAs与靶基因的结合,从而影响miRNAs对靶基因的调控作用[7]。2)参与蛋白质的形成;一些研究发现,许多circRNAs具有内部核糖体进入位点(internal ribosome entry site,IRES)或开放阅读框(open reading frame,ORF),参与功能蛋白的转录和翻译[24]。3)与蛋白质的互作作用;4)促进亲本基因的转录[9];例如,由circ-EIF3J和circ-PIAP2形成的EiciRNAs-u1 SNRNP化合物可以与RNA聚合酶Ⅱ相互作用,促进亲本基因转录过程[25]。5)作为翻译模板[26];6)作为生物标记分子;如:癌症和肿瘤[27-28]、心血管系统疾病[29]、肠道疾病等[30]。
2 circRNAs对哺乳动物肠屏障功能的影响及作用机制
2.1 circRNAs对哺乳动物肠道机械屏障的影响及作用机制
哺乳动物的肠道机械屏障由肠上皮细胞、细胞间连接复合物组成[31],其中肠道上皮细胞是肠道物理屏障的最强决定因素。肠上皮细胞由肠道隐窝中的干细胞增殖分化而来,包括吸收性肠上皮细胞、杯状细胞、肠内分泌细胞、潘氏细胞等[32]。肠道干细胞通过调控肠道上皮细胞的更新和再生进而影响肠道机械屏障功能,现有研究表明circRNAs对哺乳动物肠道干细胞及肠道上皮细胞具有调节作用并影响动物肠道屏障功能。Zhu等[33]研究发现,小鼠和人类的肠道干细胞(intestinal stem cells,ISCs)中circPan3高表达,并以一种依赖于免疫细胞的方式参与了肠干细胞的自我更新,进一步研究发现,circPan3与白细胞介素13受体α1亚基(interleukin-13 receptor subunit alpha-1,IL-13Rα1)的mRNA结合,从而促进IL-13Rα1在肠干细胞中稳定表达;同时,由Ⅱ型固有淋巴细胞(type 2 innate lymphoid cell,ILC2)分泌的白细胞介素13(interleukin 13,IL-13)与IL-13Rα1结合并激活Foxp1的表达,促进β-Catenin入核,参与肠干细胞的自我更新。Guo等[34]研究发现,在小鼠小肠隐窝和结肠中circBtnl1可负调控ISCs的自我更新,敲除circBtnl1后肠道自我更新和上皮细胞再生能力增强;进一步研究发现,circBtnl1可与DEAD解旋酶因子(DEAD-box helicase 3 Y-linked,DDX3Y)蛋白相互作用,进而影响Atf4 mRNA的稳定性,且Sox9转录被抑制,导致ISCs的自我更新能力和上皮再生减弱。以上研究表明,circRNAs可调控肠道上皮细胞的周转更新,间接调控肠道机械屏障功能。
肠道上皮细胞是维持肠道机械屏障功能的基础,现有研究表明circRNAs通过多种方式参与调控肠道上皮细胞生理状态进而影响肠道屏障功能。Li等[35]构建了体外猪小肠上皮细胞(porcine intestinal epithelial cells,IPEC-J2)氧化应激模型,研究发现circGLI3发挥“海绵作用”吸附miR-339-5p,进而调控VEGFA基因的表达,且过表达circGLI3可促进IPEC-J2的增殖并显着提高细胞中抗氧化酶活性,促进肠道机械屏障的稳态。方满新[36]研究发现,IPEC-J2感染猪圆环病毒2型(porcine circovirus type 2,PCV2)后,circRNA4410可与lncRNA MSTRG.19 762.1竞争性结合ssc-miR-429-3p进而解除ssc-miR-429-3p对靶基因IL1A的抑制作用,从而减少IPEC-J2的凋亡且使IL-8、 IL-6、TNF-α、IL-7等细胞因子表达下调。Zhao等[37]研究发现,传染性胃肠炎冠状病毒感染IPEC-J2后,细胞发生炎症反应并死亡,且circEZH2表达下调,进一步研究发现circEZH2通过“海绵化”miR-22调控circEZH2/miR-22/HK2轴和circEZH2/miR-22/IL-6/NF-κB轴,减缓细胞炎症反应。上述研究表明,circRNAs通过调控肠道上皮细胞的生理状态间接参与肠道机械屏障功能的维持。
研究发现,动物肠道黏膜受损时,circRNAs参与调节肠道机械屏障功能,使肠道维持其正常生理功能。Liu等[38]研究发现,circ_0001105在患脓毒症的大鼠肠黏膜组织表达下调,而上调circ_0001105后降低了肠道中炎症因子和Yes关联蛋白(Yes-associated protein,YAP)的表达,并缓解了肠道的氧化损伤,从而维持了患脓毒症大鼠肠道的肠道屏障功能。Ye等[39]研究发现,脓毒症患者肠道隐窝细胞中circFLNA表达上调,进一步研究发现circFLNA通过“海绵化”miR-766-3p增强了Fas介导的细胞凋亡信号通路,且circFLNA在体内外模型中均通过miR-766-3p/Fas轴促进肠上皮细胞凋亡和炎症反应。Liu等[40]研究发现,在患脓毒症大鼠中沉默circDNMT3B会加剧肠组织氧化损伤、增加炎症因子水平,miR-20b-5p可上调脓毒症大鼠肠道中炎症因子水平、破坏肠道黏膜屏障,进一步研究发现circDNMT3B可作为miR-20b-5p“海绵”,进而负向调节miR-20b-5p对肠道黏膜屏障的破坏作用。Zhang等[41]发现,在严重烧伤导致肠黏膜受损的小鼠中过表达circRNA_Maml2可促进肠上皮细胞的增殖和迁移,其具体机制为circRNA_Maml2可作为“miRNA海绵”结合miR-93-3p,激活FZD7介导的Wnt/β-catenin信号通路,促进肠道上皮细胞增殖和迁移,加快严重烧伤后受损肠黏膜屏障的修复。Deng等[42]也在严重烧伤小鼠肠道中发现circMaml2的表达明显降低,而过表达circMaml2可改善严重烧伤后的肠黏膜损伤和促进肠道屏障的修复,进一步研究表明在小鼠肠上皮细胞中circMaml2作为“miRNA海绵”与miR-683结合,调控miR-683对靶基因Sec62的作用,同时通过PTBP1通路促进受损肠黏膜的重建。Yang等[43]研究发现,N-草酰基-D-苯丙氨酸(N-oxalyl-D-phenylalanine,NOFD)可增强HIF-1α(hypoxia-inducible factor,HIF)的转录活性,减轻电离辐射造成的胃肠道损伤,维持肠上皮屏障功能;并且发现NOFD可促进circRNA_2909和circRNA_0323与mmu-miR- 92a-1-5p竞争性结合,解除对HIF-1α基因的抑制,使HIF-1α的转录活性增强,促进胃肠道损伤修复。营养物质也可通过影响circRNAs表达参与肠道屏障功能调控。Li等[44]研究发现,香叶木素可缓解由葡聚糖硫酸钠(dextran sulfate sodium,DDS)导致肠道屏障功能的破坏,进一步研究发现香叶木素增加紧密连接蛋白的表达并降低肠道中炎症因子水平,还可激活circ-Sirt1/Sirt1轴抑制NF-κB通路减缓由DDS导致的小鼠结肠氧化应激和炎症反应,并促进肠道屏障的稳态。
细胞间连接复合物中的紧密连接是由跨膜蛋白(如:claudins,occluddin)、外周膜蛋白(如:zonula occluddens (ZO)-1,ZO-2,ZO-3)和调节蛋白相互连接形成的封闭复合物,是维持肠道上皮屏障功能和控制肠道上皮通透性的重要因素[45]。现有研究表明,circRNAs可调控跨膜蛋白及外周膜蛋白的表达影响肠道屏障功能。Rankin等[46]研究发现,长链非编码RNA CDKN2B-AS1存在两种表达形式——线型和环型,且两种形式在炎症性肠病患者的结肠细胞中均显着下调,CDKN2B-AS1下调后结肠细胞数量显着增多。此外,CDKN2B-AS1的下调导致紧密连接蛋白Claudin-2也下调,从而增强了结肠细胞的屏障功能。Zhao等[47]研究发现,在克罗恩患者结肠组织中circSMAD4表达显着上调,且过表达circSMAD4显着下调occludin和ZO-1的表达,促进上皮细胞的凋亡,进一步研究发现circSMAD4通过与miR-135a-5p相互作用促进JAK2激酶表达,导致肠道屏障破坏并促进克罗恩结肠炎的发展。
虽然许多circRNAs在肠道黏膜机械屏障中的作用机制仍然不是十分清楚,但根据以上研究表明,肠道屏障受损后circRNAs参与修复过程并对肠道稳态的维持发挥重要调控作用。
2.2 circRNAs对哺乳动物肠道化学屏障的影响及作用机制
肠道化学屏障由胃酸、抗菌肽、溶菌酶等化学物质构成。潘氏细胞分泌的肽类和蛋白在抵御外来致病菌及维持肠道化学屏障功能稳态等方面具有重要作用。现有研究表明,circRNAs可通过调控潘氏细胞功能影响肠道化学屏障。Xiao等[48]建立盲肠结扎和穿刺诱导的小鼠损伤模型进行研究,发现在肠上皮稳态和肠道黏膜病理状态发生变化时,circ-HIPK3的表达水平会产生显着性差异,并伴有潘氏细胞缺陷、损伤,虽然circHIPK3在潘氏细胞中所发挥的调控作用尚不清楚,但这些发现表明,circ-HIPK3在肠道化学屏障中具有调节作用。Li等[49]研究发现,在肠道黏膜损伤或炎症性肠病患者肠上皮细胞中HuR和ATG16L1基因表达下降且潘氏细胞数量减少,已有的研究表明ATG16L1基因表达会导致潘氏细胞损伤,进一步研究发现circPABPN1可抑制HuR与ATG16L1 mRNA的结合,进而抑制ATG16L1的翻译,从而参与调节肠道潘氏细胞功能。
在miRNAs的研究中也发现,miRNAs对肠道化学屏障具有调节作用,如miR-195过表达小鼠的肠道黏膜组织中簇细胞和潘氏细胞减少,但对杯状细胞和肠道上皮细胞无影响,其机制为miR-195与HuR蛋白竞争性结合Dclk1 mRNA从而抑制DclkmRNA的翻译,损坏了簇细胞和潘氏细胞的功能[50]。Zhu[51]等研究发现,hsa_circ_0013401作为miR-195的“海绵”下调PAK2基因的表达从而调节神经母细胞瘤的生理活动。由此推断,miR-195在肠道化学屏障中的作用受到circRNAs调控。Mo[52]等研究发现,miR-429在小鼠结肠炎组织中表达下调,而MARCKS基因表达水平上调,MARCKS可调控肠道粘蛋白2的分泌,进一步研究发现miR-429可与MARCKS靶向结合共同调控肠道化学屏障。Li[53]等研究发现,circPPARA可靶向miR-429和miR-200b调控猪肌肉内前脂肪细胞的分化和增殖,影响猪肌肉内脂肪的生成。由上述研究可知,circRNAs可通过介导miRNAs的表达在哺乳动物化学屏障中的产生调控作用。
circRNAs在动物肠道化学屏障中调节机制的研究较少,但以上研究表明circRNAs可调节化学屏障细胞的生理状态参与肠道化学屏障调控。
2.3 circRNAs对哺乳动物肠道免疫屏障的影响及作用机制
先天性淋巴细胞(innate lymphoid cells,ILCs)是一类免疫效应细胞,在宿主防御、代谢稳态和组织修复等方面具有重要作用,先天性淋巴细胞在细胞因子介导的肠上皮细胞屏障完整性调控中发挥着重要作用。3型先天淋巴样细胞(group 3 innate lymphoid cell,ILC3)是ILCs的亚群之一[54],ILC3在肠道免疫、炎症和组织稳态中起调节作用,现有研究证实circRNAs可调节ILC3功能进而影响肠道免疫屏障。Liu等[55]研究发现,用葡聚糖硫酸钠(dextran sodium sulfate,DSS)处理小鼠后观察到ILC3的激活被抑制,小鼠肠道3型先天淋巴样细胞中circKcnt2的表达显着上调,进一步研究发现circ-Kcnt2与核小体重构脱乙酰酶(nuclesome remodeling deactylase,NuRD)复合物中的Mbd3相互作用,将NuRD复合物招募到ILC3中的Batf启动子上,抑制Batf转录,从而抑制ILC3的激活,促进先天结肠炎的愈合。Liu等[56]研究发现,在ILC3中circZbtb20高表达,进一步研究发现circZbtb20可增强Alkbh5和Nr4a1 mRNA之间的相互作用并促进Nr4a1 mRNA稳定性,Nr4a1启动Notch2信号通路的激活,这有助于维持ILC3的功能,从而维持了肠道的稳态。Liu等[57]研究发现,circTmem241在ILC3及先天淋巴细胞祖细胞中高表达,且在缺失circTmem241后ILC3s的抗菌免疫功能被减弱;在先天淋巴前体细胞(innate lymphoid cell precursors,ILCPs)中,circ-Tmem241能够与Nono蛋白相互作用,从而招募组蛋白甲基转移酶 Ash1l到Elk3的启动子上,促进Elk3转录,并最终影响ILC3s的分化及其抗菌免疫功能。除此之外,有研究发现circRNAs直接参与细胞免疫应答反应基因,调控肠道免疫屏障。OuYang等[58]研究发现,circ_0001187在结肠炎患者结肠黏膜细胞及血清外泌体中过表达,通过利用肿瘤坏死因子-α刺激人正常结直肠黏膜细胞(human normal colorectal mucosa cells,FHC) 构建细胞炎症模型进行探究,发现下调circ_0001187可促进FHC的增殖,并抑制FHC炎症反应和氧化应激,其具体机制为circ_0001187可以“海绵化”miR-1236-3p,导致miR-1236-3p的靶基因髓系分化因子88(myeloid differentiation factor 88,MYD88)表达下调,MYD88是细胞免疫应答反应中的关键基因。
此外,在生物医学领域RNA核酸疫苗的研发也揭示了cirRNAs参与机体免疫活动的调控。2022年,Qu等[59]成功研发针对新型冠状病毒的环状RNA疫苗,研究表明该环状 RNA 疫苗在小鼠和恒河猴体内可以有效降低新冠病毒感染的恒河猴肺部的病毒载量,显着缓解新冠病毒感染引起的肺炎症状。同年,Li等[60]建立了一种新的circRNA疫苗平台,该疫苗能够触发机体强大的先天和适应性免疫激活,并在多种小鼠肿瘤模型中显示出优越的抗肿瘤疗效。上述研究为畜牧生产中畜禽免疫治疗提供了一定研究思路。
上述研究表明,circRNAs参与动物肠道屏障功能调控,表1总结了circRNAs在肠道机械屏障、化学屏障及免疫屏障中的作用机制研究进展。
2.4 circRNAs对哺乳动物肠道生物屏障的影响及作用机制
哺乳动物肠道中存在大量的微生物,这些微生物依赖动物肠道生存,它们也是肠道生物屏障的重要组成部分。研究发现,circRNAs与肠道微生物可相互调控并影响大脑的生理功能。Chen等[61]研究发现,高糖高脂饮食会使小鼠肠道菌群失调及导致小鼠大脑circRNAs表达产生差异;且这些差异表达的circRNAs(如circNF1-419和circ_0001239)参与了小鼠肠道微生物的调控。该团队还对快速老化(SAMP8)小鼠进行了研究,发现在SAMP8小鼠大脑皮层中过表达circNF1-419,可以改善肠道组织的损伤并改变小鼠肠道微生物组成[62]。肠道微生物也可调控circRNAs的表达参与多种生理过程。Zhu等[63]研究发现,广谱抗生素(antibiotics,ABX)能够促进癌症转移,而将无特定病原体(specific pathogen-free,SPF)小鼠的粪便微生物群移植或将双歧杆菌灌胃注入无菌小鼠或ABX小鼠中,可显着抑制肿瘤转移;进一步研究发现,肠道微生物群以IL-11依赖的方式抑制mmu_circ_0000730的表达,mmu_circ_0000730通过与mmu-miR-466i-3p、mmumiR-466 f-3p相互作用,间接调控靶基因SOX9表达。由此可知,肠道微生物群通过IL-11/circRNA/miRNA轴参与癌症转移调控。Guo等[64]研究发现,冷暖季节藏绵羊瘤胃上皮细胞中circRNAs表达存在差异,这些差异表达circRNAs参与调控氨基酸和蛋白质合成及短链脂肪酸和挥发性脂肪酸转运;circRNAs还参与调控瘤胃微生物区系,促进瘤胃微生物定植,使藏羊适应寒冷季节的营养胁迫。
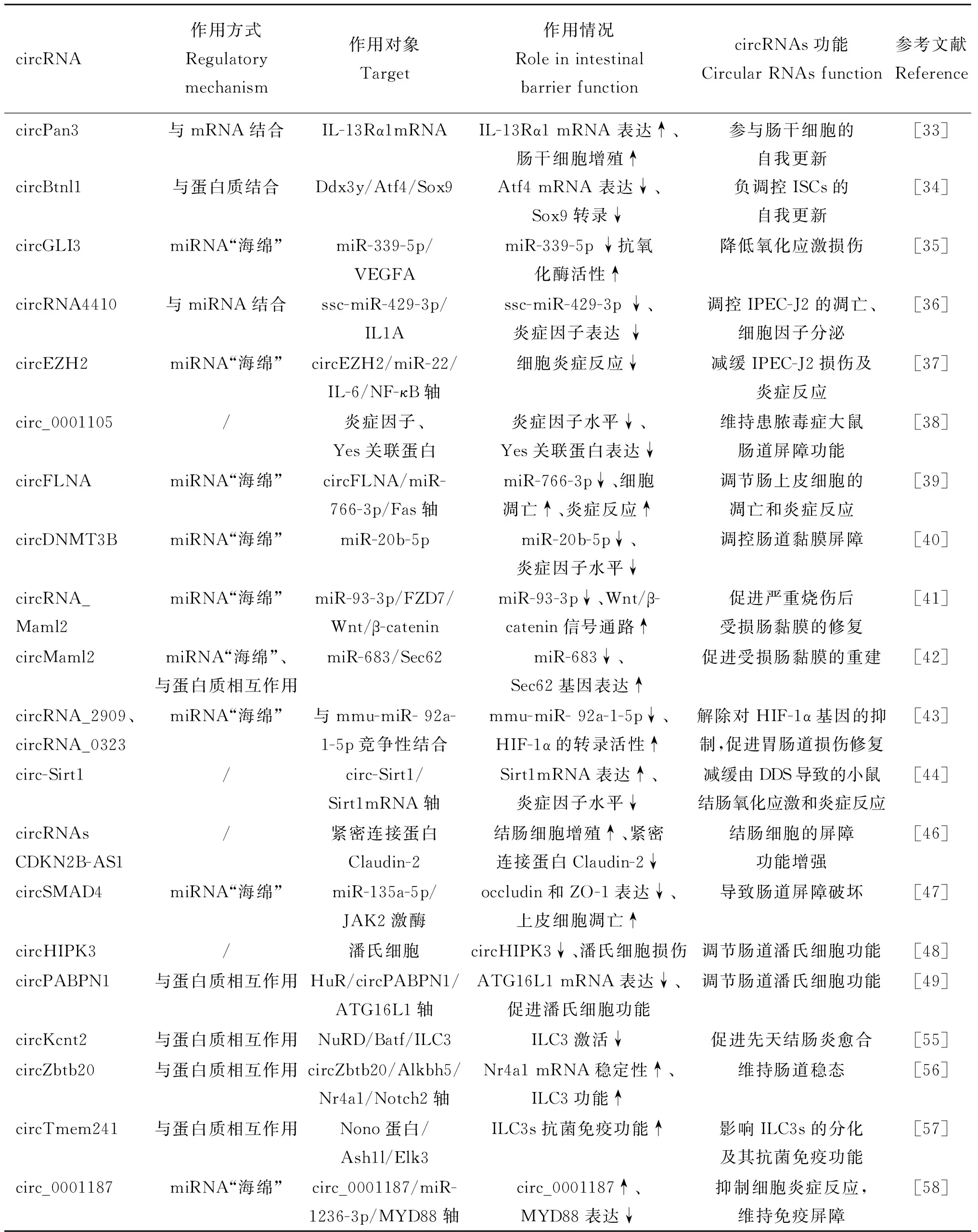
表1 circRNAs在哺乳动物肠道屏障功能中的作用Table 1 The role of circRNAs in mammalian intestinal barrier function
目前,circRNAs在动物肠道微生物屏障中的作用机制研究较少,但上述研究表明circRNAs对肠道微生物之间存在调节作用,并且该作用可通过脑-肠轴影响大脑的生理状态。
3 展 望
随着高通量测序技术和生物信息学技术的发展,被认为是转录“噪音”的circRNAs的研究热度不断上升,circRNAs在多种生理活动中所发挥的作用也逐渐探明,其为探究哺乳动物肠道屏障调控机制提供了新路径。目前的研究表明,circRNAs在哺乳动物肠道机械屏障和肠道免疫屏障中主要是通过发挥“miRNAs海绵”作用及蛋白质互作作用参与肠道屏障功能调控;circRNAs在肠道化学屏障中的作用机制研究较少,根据目前的研究推测circRNAs对肠道化学屏障的调控作用主要是通过miRNA实现的,但其具体机制还有待探究;circRNAs在肠道微生物屏障中的作用机制暂不明确。此外,研究表明香叶木素等可影响circRNAs的表达进而调控肠道屏障生理功能。在畜牧生产上,未来的研究应重点关注如何利用营养素调控circRNAs的表达进而调控畜禽肠道健康,提高畜禽生产效率,节约生产成本。



