刘芳 徐细明

[摘要] 目的 探讨细胞周期相关因子3(CDCA3)在肝癌中的生物学行为。 方法 收集2018年1月~2019年6月在武汉大学人民医院肝胆外科行手术治疗的肝细胞肝癌患者的组织标本,共30例肝癌标本和癌旁标本;分别采用Real-Time PCR检测不同组织中的CDCA3的表达情况;采用lip3000转染法分别将CDCA3干扰RNA序列和对照序列转入肝癌细胞株,应用CCK-8、EdU免疫荧光染色检测细胞增殖,流逝细胞术检测细胞周期。 结果 肝癌组织CDCA3的表达量明显高于癌旁组织,差异有高度统计学意义(P < 0.01)。HepG2、Huh7、SMMC-7721细胞中CDCA3的表达量明显高于LO2细胞,差异有统计学意义(P < 0.05或P < 0.01),而BEL-7402细胞中CDCA3的表达量与LO2细胞的表达量比较,差异无统计学意义(P > 0.05)。siCDCA3#1和siCDCA3#2下调SMMC-7721细胞CDCA3的表达量与对照组(siNC)比较,差异均有统计学意义(均P < 0.05),故选取SMMC-7721细胞完成后续研究。siCDCA3#1和siCDCA3#2下调SMMC-7721细胞CDCA3的表达量后,24 h和48 h细胞增殖数明显低于siNC,差异均有统计学意义(均P < 0.05)。并且EdU检测阳性细胞数明显低于siNC。siCDCA3#2下调SMMC-7721细胞CDCA3表达量后,细胞周期的G1期时间较siNC延长,差异有统计学意义(P < 0.05)。 结论 CDCA3能够促进肝癌细胞增殖,是潜在的治疗靶点。
[关键词] 细胞周期相关因子3;肝癌;增殖;细胞周期
[中图分类号] R735.7 [文献标识码] A [文章编号] 1673-7210(2020)05(b)-0014-06
Study on the role of CDCA3 in the proliferation of hepatocellular carcinoma
LIU Fang XU Ximing
Cancer Center, Renmin Hospital of Wuhan University, Hubei Province, Wuhan 430060, China
[Abstract] Objective To investigate the biological behavior of cell division cycle associated protein 3 (CDCA3) in liver cancer. Methods Tissue specimens of hepatocellular carcinoma patients who underwent surgical treatment in the Department of Hepatobiliary Surgery, Renmin Hospital of Wuhan University from January 2018 to June 2019 were collected. There were 30 liver cancer specimens and paracancer specimens. Real-time PCR was used to detect the expression of CDCA3 in different tissues. CDCA3 interfering RNA sequences and control sequences were transfected into hepatocellular carcinoma cell lines by lip3000 transfection. Cell proliferation was detected by CCK-8 and EdU immunofluorescence staining, and cell cycle was detected by elapse cytology. Results The expression level of CDCA3 in liver cancer tissues was significantly higher than that in paracancer tissues, and the difference was highly statistically significant (P < 0.01). The expression levels of CDCA3 in HepG2, Huh7 and SMMC-7721 cells were significantly higher than those in LO2 cells, with statistically significant differences (P < 0.05 or P < 0.01), while the expression level of CDCA3 in BEL-7402 cells was not statistically significant compared with that in LO2 cells (P > 0.05). The expression of CDCA3 in SMMC-7721 cells down-regulated by SiCDCA3#1 and siCDCA3#2 was statistically significant compared with that in the control group (siNC) (P < 0.05). Therefore, SMMC-7721 cells were selected to complete the follow-up study. After the expression levels of CDCA3 in SMMC-7721 cells were down-regulated by siCDCA3#1 and siCDCA3#2, the number of cell proliferation at 24 h and 48 h was significantly lower than that in the siNC, and the differences were statistically significant (all P < 0.05). And the number of positive cells detected at EdU was significantly lower than that in the siNC. Compared with the siNC, the G1 phase time of SMMC-7721 cell cycle was prolonged after the expression of CDCA3 in SMMC-7721 cell was down-regulated by siCDCA3#2, and the difference was statistically significant (P < 0.05). Conclusion CDCA3 can promote the proliferation of liver cancer cells and is a potential therapeutic target.
[Key words] Cell division cycle associated protein 3; Liver cancer; Proliferation; Cell cycle
原发性肝癌(以下简称“肝癌”)是临床常见的恶性肿瘤,位居世界癌症死亡率第5位,5年生存率约为18%[1]。早期肝癌患者的治疗主要以手术或局部消融为主,中晚期患者则多采用介入、放化疗或靶向等手段[2-3]。受限于当前缺少足够特异和有效的手段针对高危人群进行早期诊断和临床前筛查,因此,当肝癌确诊时肿瘤已演进多时,导致仅有约1/3的患者适合手术切除。以上种种因素导致了肝癌患者往往容易复发及转移,5年生存率仍处于较低水平[4]。鉴于当前所面临的种种困境,寻找肝癌相关标志物,研究其调控的分子机制,具有重要的理论和临床意义。
前期的研究中,笔者团队通过基因芯片技术在肝癌组织和癌旁组织中筛选出具有差异表达的信使RNA(mRNA)[5-6]。基因芯片数据表明肝癌组织中细胞周期相关因子3(cell division cycle associated protein 3,CDCA3)的mRNA表达水平较正常肝组织高。CDCA3,又称Tome-1,是细胞进行有丝分裂过程中所必需的胞质蛋白,其调控着细胞周期的进展[7]。它可以通过淘汰素、S期激酶相关蛋白L共同参与E3连接酶复合物的构成,与丝氨酸/苏氨酸蛋白激酶、有丝分裂抑制激酶家族成员WEEL的泛素化和降解相关,它是细胞进入有丝分裂的触发器[8],同时CDCA3也参与细胞周期G1期的调控[9]。近年来,研究发现CDCA3在非小细胞肺癌[7]、胃癌[10]、膀胱癌[11]、白血病[12]、结肠癌[13]、乳腺癌[9]等肿瘤中出现异常高表达的现象,表明其可能在肿瘤的发生发展中起到至关重要的作用,但是在肝癌中的研究较少。本研究旨在探讨CDCA3在肝癌中的作用,为肝癌的诊断及治疗提供新的分子靶标。
1 资料与方法
1.1 一般资料
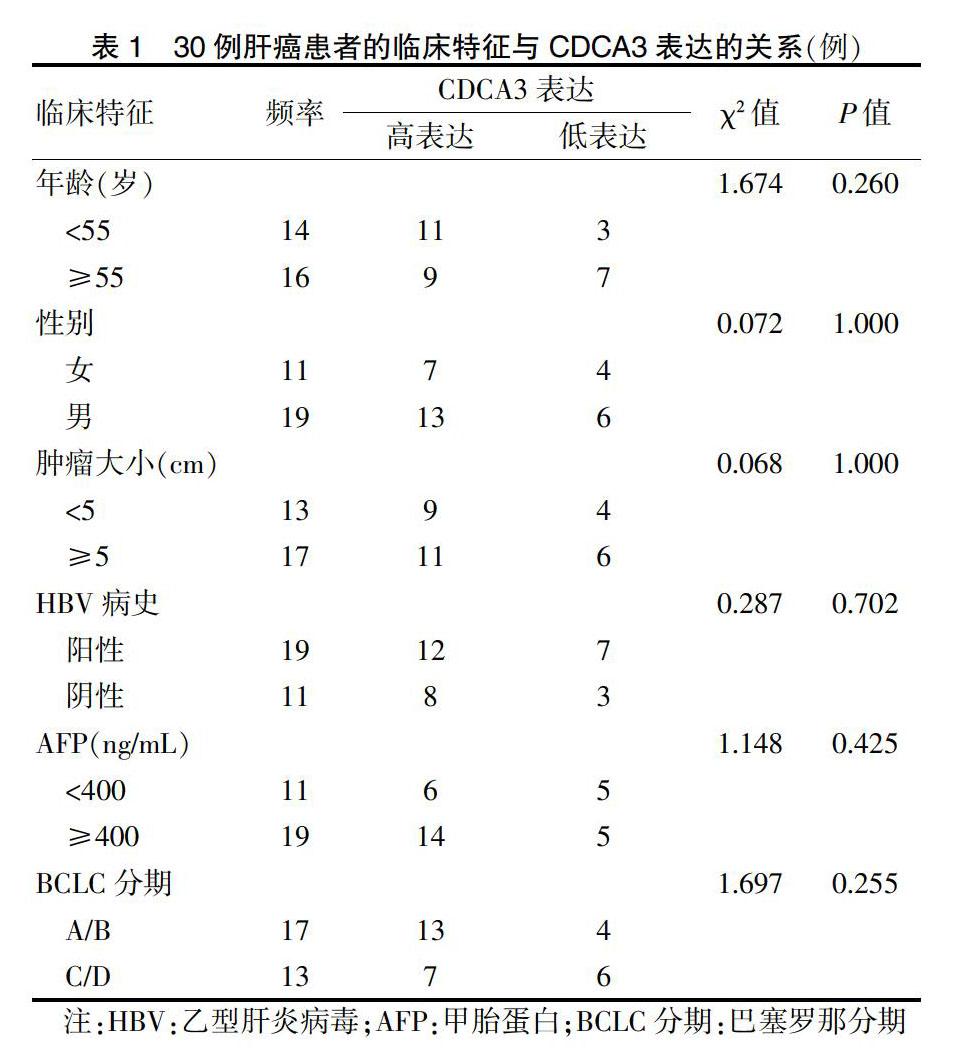
所选30对标本均来自于武汉大学人民医院(以下简称“我院”)肝胆外科2018年1月~2019年6月行肝癌切除术患者所切除的肝癌组织及癌旁组织(距离手术切缘至少2 cm)。术前患者均未接受放化疗、射频消融等辅助治疗,术后病理结果证实为肝细胞肝癌,患者的临床特征如表1所示。所有标本装入冻存管后迅速保存于液氮中。所有标本的获取均经过患者允许,本研究经我院医学伦理委员会审批。
1.2 细胞培养
正常肝细胞系LO2、人肝癌细胞系HepG2、SMMC-7721、Huh7和BEL-7402购买于上海细胞生物学研究所,HepG2、SMMC-7721和BEL-7402采用含有10%胎牛血清(Gibico,南美)和1%青-链霉素(谷歌生物,中国)的高糖培养基(Hyclone,美国),正常肝细胞系LO2和Huh7采用含有10%胎牛血清(Gibico,南美)和1%青-链霉素(谷歌生物,中国)的高糖培养基(Hyclone,美国)置于37℃、5% CO2细胞培养箱培养(Thermo,美国)。按照70%高糖培养基,20%血清和10%二甲基亚砜(Dimethylsulfoxide,DMSO)进行梯度冻存,保存于液氮。
1.3 细胞瞬时转染
将生长良好处于对数生长期的肝癌细胞接种于6孔板中,当细胞融合度在70%~80%,弃培养基,换成无血清和抗生素的培养基,置于恒温培养箱中培养。采用lip3000(L3000015,吉凯基因,中国)脂质体转染的方法,将干扰RNA(small interfering RNA,siRNA)或者对照序列(S151029151613,锐博生物,中国)用无血清培养基按照说明进行稀释,室温静置5 min后,再轻轻地加入稀释的lip3000,室温孵育20 min后轻轻加入6孔板对应的孔中。转染6~12 h后,换用完全培养基继续培养,按照实验所需时间收集转染细胞。
1.4 实时定量聚合酶链反应(Real Time-PCR)
细胞系和组织标本处理:用Trizol(C10268951,Invitrogen,美国)分别提取正常肝细胞、肝癌细胞、30例原发性肝癌组织和对应癌旁组织的总RNA,然后按TOYOBO试剂盒(QPK-201,上海基星生物科技有限公司,日本)说明书将RNA反转录成cDNA。
干扰CDCA3表达样品处理:将CDCA3的siRNA及对照序列分别转染到肝癌细胞系。转染48 h后,用Trizol提取总RNA,并用Nano-Drop2000C紫外分光光度计(Thermo,美国)测定RNA浓度及纯度。然后按TOYOBO试剂盒(QPK-201,上海基星生物科技有限公司,日本)说明书将RNA反转录成cDNA。
最后按照Takara qRT-PCR试剂盒说明书进行操作,20 μL体系,设立3个复孔,甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GDPH)为内参基因,7500 Fast System SDS软件(BIO-RAD,美国)分析Ct值,按95 ℃ 10 min,90 ℃ 10 s,56 ℃ 30 s,72 ℃ 32 s,40个循环进行扩增,采用2-△△Ct法计算相对表达量。引物序列由武汉巴菲尔生物有限公司设计合成。见表2。
1.5 细胞计数(CCK-8)检测肝癌细胞增殖
选取处于指数生长期的细胞,精确计数后将细胞调整为(3~4)×104个/mL接种于96孔板,待细胞贴壁后,对细胞进行干预实验,设置3个重复孔。分别于0、6、24、48 h,弃培养基,加入CCK-8混合反应液,37℃孵育2 h后用酶标仪检测各孔450 nm的吸光度值。
1.6 EdU免疫荧光检测肝癌细胞增殖
选取(3~4)×104个/mL的肝癌细胞接种于96孔板中,待细胞贴壁后对细胞进行干预实验,设置3个重复孔。细胞干预48 h后,弃培养基,加入按照1000∶1的比例稀释EdU溶液(CA1170,锐博生物,中国)孵育2 h,弃培养液,用磷酸缓冲盐溶液(phosphate buffer saline,PBS)(Hyclone,美国)清洗2遍。每孔中加入50 μL 4%多聚甲醛(谷歌生物,中国)室温孵育20 min后,弃废液;每孔加入50 μL甘氨酸溶液(CA1170,锐博生物,中国),清洗5 min,弃废液;用PBS清洗5 min,弃废液;每孔加入100 μL渗透剂(0.5% Tritonx-100的PBS)脱色摇床孵育10 min,PBS清洗5 min,弃废液;每孔中加入100 μL的1×Apollo染色反应液(CA1170,锐博生物,中国),室温下避光,置于脱色摇床上孵育30 min,弃废液;加入100 μL渗透剂脱色摇床清洗2~3次,每次5 min,PBS清洗5 min;每孔加入100 μL 1×Hochest反应液,脱色摇床室温避光孵育30 min,弃废液,PBS清洗3次。用倒置荧光显微镜(Olympus,日本)观察并拍照。
1.7 流式细胞术检测肝癌细胞周期检测
收集转染48 h后的细胞,清洗离心后用固定液(1×)配成1×106个/mL的细胞悬液,再加入10 μL试剂B(70-APCC101,联科生物,中国),室温避光孵育30 min。用流式细胞仪分析(Becton-Dickinson,USA),采用FACS express version 3软件分析细胞周期变化情况。
1.8 统计学方法
采用IBM SPSS 20.0(IBM,USA)对所得数据进行统计学分析,计量资料采用均数±标准差(x±s)表示,两组比较采用t检验,多组样本间比较用ANOVA检验;计数资料采用百分率表示,采用χ2检验或Fisher确切概率法。以P < 0.05为差异有统计学意义。所有的图表运用GraphPad Prism 5(GraphPad Software,CA,USA)制作。
2 结果
2.1 CDCA3在肝癌标本中表达情况
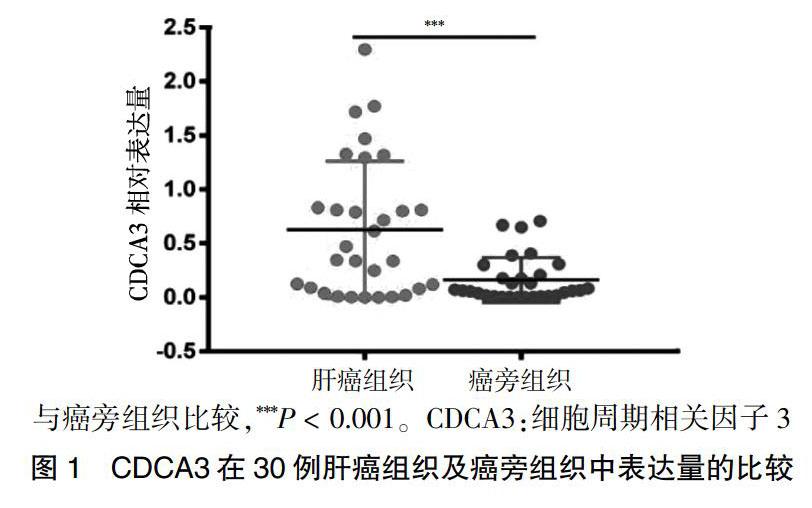
肝癌组织CDCA3的表达量明显高于癌旁组织的表达量,差异有高度统计学意义(P < 0.01)。见图1。进一步分析患者信息发现CDCA3表达量高低与患者年龄、性别、肿瘤大小、HBV病史、AFP和BCLC分期无相关性(P > 0.05)。见图1。
2.2 CDCA3在肝癌细胞系中表达情况
HepG2、Huh7、SMMC-7721细胞中CDCA3的表达量明显高于LO2细胞的表达量,差异有统计学意义(P < 0.05或P < 0.01)),而BEL-7402细胞中CDCA3的表达量与LO2细胞的表达量比较,差异无统计学意义(P > 0.05)。见图2。故选取HepG2、SMMC-7721和Huh7细胞作为后续实验的肝癌细胞系。
2.3 通过siRNA下调CDCA3的表达量
siCDCA3#1和siCDCA3#2下调SMMC-7721细胞CDCA3的表达量与对照组(siNC)比较,差异均有统计学意义(均P < 0.05);siCDCA3#1下调Huh7细胞CDCA3的表达量与对照组(siNC)比较,差异有统计学意义(P < 0.05),而siCDCA3#2下调Huh7细胞CDCA3的表达量与对照组(siNC)比较,差异无统计学意义(P > 0.05);siCDCA3#1和siCDCA3#2下调HepG2细胞CDCA3的表达量与对照组(siNC)比较,差异均无统计学意义(均P > 0.05),见图3。故选取SMMC-7721作为后期研究的肝癌细胞系。
与对照组(siNC)比较,*P < 0.05。CDCA3:细胞周期相关因子3
图3 不同siRNA序列与对照序列siNC分别转染肝癌细胞后CDCA3表达量的比较
2.4 CCK-8检测下调CDCA3表达量对肝癌细胞增殖的影响
siCDCA3#1和siCDCA3#2下调SMMC-7721细胞CDCA3的表达量后,24 h和48 h细胞增殖数明显低于对照组(siNC),差异均有统计学意义(均P < 0.05)。见图4。
不同时间段细胞增殖数与对照组(siNC)比较
2.5 EdU检测干扰CDCA3表达对肝癌细胞增殖的影响
siCDCA3#1和siCDCA3#2下调SMMC-7721细胞CDCA3的表达量后,阳性细胞数(红色)明显低于对照组(siNC)。见图5(封四)。
2.6 流式细胞仪检测下调CDCA3对肝癌细胞周期的影响
与对照组(siNC)比较,siCDCA3#2下调SMMC-7721细胞CDCA3表达量后,细胞周期的G1期时间延长,差异有统计学意义(P < 0.05),细胞周期的S、G2期时间差异无统计学意义(P > 0.05)。而siCDCA3#1下调SMMC-7721细胞CDCA3表达量后,细胞周期的G1、S、G2期时间与对照组(siNC)比较,差异无统计学意义(P > 0.05)。见图6。
3 讨论
尽管分子诊断和治疗策略快速发展,但目前原发性肝癌没有特异性的治疗靶点。因此,急需进一步研究发现更有效的肝癌生物标志物。研究表明细胞周期调节基因的异常表达可导致肿瘤发生,推测细胞周期调节蛋白抑制剂可能成为治疗肿瘤的新疗法[14]。在前期研究中,本课题组通过高通量测序发现CDCA3在肝癌组织中显着高表达[5-6],CDCA3是细胞进入有丝分裂的触发器,且是细胞周期蛋白依赖激酶1(cdk1)/细胞周期蛋白B(cyclin B)的适当激活和细胞进入有丝分裂所必需的蛋白质[15-16]。CDCA3编码蛋白含有286个细胞有丝分裂的必需胞质蛋白,通过控制有丝分裂入口从而来调节细胞周期[8]。细胞周期的异常调控将导致细胞的过度增殖失控,促使恶性肿瘤的形成,其在肿瘤的发生发展中具有重要意义。CDCA3可以通过影响DNA甲基化,从影响细胞周期的进展[17]。
为了进一步验证CDCA3在肝癌中的作用,Real Time-PCR检测发现CDCA3在30例原发性肝癌组织中高表达,与前期高通量测序结果吻合[6]。但由于随访时间太短,本研究尚未确定CDCA3表达水平与患者生存时间的相关性。通过细胞实验,发现下调CDCA3可以抑制肝癌细胞增殖和细胞周期进展。Hu等[18]发现CDCA3可能协同OY-TES-1共同参与肝癌细胞的增殖、迁移、侵袭和凋亡,与本研究结果相符。因此,本研究推测CDCA3可能是原发性肝癌的潜在生物标志物。
根据最近的研究,CDCA3可以通过激活NF-κB/cyclin D1信号通路,从而影响肿瘤细胞增殖,导致肿瘤的发生[17]。CDCA3是否能通过NF-κB/cyclin D1信号通路影响肝癌细胞增殖,后续研究会进一步证实。当然本研究也有不足之处:①只检查了30例原发性肝癌的组织标本,可能不具有代表性;②细胞系太过单一,应选多株细胞进行实验;③只是单从细胞水平进行了验证,需要进一步行动物实验来验证CDCA3在原发性肝癌中的功能。总体而言,进一步的研究需要收集足够的样品,并且建立动物模型,探讨CDCA3与NF-κB/cyclin D1信号通路的关系。
综上所述,本研究提示,CDCA3可能是癌症进展的关键分子,是预防和治疗肝癌的潜在生物标志物和治疗靶点。
[参考文献]
[1] Siegel RL,Miller KD,Jemal A. Cancer statistics,2019 [J]. CA Cancer J Clin,2019,69(1):7-34.
[2] Song T. Recent advances in surgical treatment of hepatocellular carcinoma [J]. Drug Discov Ther,2015,9(5):319-330.
[3] Meng M,Wang H,Zeng X,et al. Stereotactic body radiation therapy:A novel treatment modality for inoperable hepatocellular carcinoma [J]. Drug Discov Ther,2015,9(5):372-379.
[4] Yang N,Li S,Li G,et al. The role of extracellular vesicles in mediating progression,metastasis and potential treatment of hepatocellular carcinoma[J]. Oncotarget,2017,8(2):3683-3695.
[5] Zhang MX,Xu XM,Zhang P,et al. Effect of silencing NEK2 on biological behaviors of HepG2 in human hepatoma cells and MAPK signal pathway [J]. Tumor Biology,2016,37(2):2023-2035.
[6] Yu TT,Xu XM,Hu Y,et al. Long noncoding RNAs in hepatitis B virus-related hepatocellular carcinoma [J]. World J Gastroenterol,2015,21(23):7208-7217.
[7] Adams MN,Burgess JT,He Y,et al. Expression of CDCA3 Is a Prognostic Biomarker and Potential Therapeutic Target in Non-Small Cell Lung Cancer [J]. J Thorac Oncol. 2017,7(12): 1071-1084.
[8] Yu J,Hua R,Zhang Y,et al. DNA hypomethylation promotes invasion and metastasis of gastric cancer cells by regulating the binding of SP1 to the CDCA3 promoter [J]. J Cell Biochem,2020,121(1):142-151.
[9] Phan NN,Wang CY,Li KL,et al. Distinct expression of CDCA3,CDCA5,and CDCA8 leads to shorter relapse free survival in breast cancer patient [J]. Oncotarget,2018,9(6):6977-6992.
[10] Zhang Y,Yin W,Cao W,et al. CDCA3 is a potential prognostic marker that promotes cell proliferation in gastric cancer [J]. Oncol Rep,2019,41(4):2471-2481.
[11] Li S,Liu X,Liu T,et al. Identification of biomarkers correlated with the TNM staging and overall survival bladder cancer [J]. Front Physiol. 2017,8:947.
[12] Bi L,Zhou B,Li H,et al. A novel miR-375-HOXB3-CDCA3/DNMT3B regulatory circuitry contributes to leukemogenesis in acute myeloid leukemia [J]. BMC Cancer,2018,18(1):182.
[13] Qian W,Zhang Z,Peng W,et al. CDCA3 mediates p21-dependent proliferation by regulating E2F1 expression in colorectal cancer [J]. Int J Oncol,2018,53(5):2021-2033.
[14] Matera R,Saif MW. New therapeutic directions for advanced pancreatic cancer: cell cycle inhibitors,stromal modifiers and conjugated therapies [J]. Expert Opin Emerg Drugs,2017,22(3):223-233.
[15] Itzel T,Scholz P,Maass T,et al. Translating bioinformatics in oncology: guilt-by-profiling analysis and identification of KIF18B and CDCA3 as novel driver genes in carcinogenesis [J]. Bioinformatics,2015,31(2):216-224.
[16] Chen J,Zhu S,Jiang N,et al. HoxB3 promotes prostate cancer cell progression by transactivating CDCA3 [J]. Cancer Lett,2013,330(2):217-224.
[17] Zhang W,Lu Y,Li X,et al. CDCA3 promotes cell proliferation by activating the NF-κB/cyclin D1 signaling pathway in colorectal cancer [J]. Biochem Biophys Res Commun.,2018,500(2):196-203.
[18] Hu Q,Fu J,Luo B,et al. OY-TES-1 may regulate the malignant behavior of liver cancer via NANOG,CD9,CCND2 and CDCA3:a bioinformatic analysis combine with RNAi and oligonucleotide microarray [J]. Oncol Rep,2015,33(4):1965-1975.
(收稿日期:2019-11-26 本文编辑:顾家毓)



