车开萱 李秦陇 杨俊超 邱俊强,2
1 北京体育大学运动人体科学学院(北京 100084)
2 运动营养北京市高等学校工程研究中心(北京 100084)
对运动训练的适应性反应是由训练时间、强度、类型和频率等多种因素决定,此外还取决于运动前后的营养质量和数量。由运动引发的适应可以通过营养信号放大或减弱。尽管以往人们长期坚信单一、优质的“运动饮食”,但运动营养实践大都基于不同运动代谢需求的特殊性以及训练和比赛目标的周期性。随着现代运动营养学的发展,运动饮食所包含的饮食方式正在以多样性、特殊性、周期化和个性化的理念支持着训练和比赛的需求[1]。
竞技运动的代谢需求是复杂的,所涉及的运动项目的持续时间从数秒(跳跃、投掷和举重)到数周(环法自行车赛)不等,最佳运动表现的实现旨在通过专项训练最大限度地激发个体遗传潜力[2]。尽管一些优秀运动员受益于系统、科学的训练和比赛建议,但结合营养策略将有助于促成他们达到运动表现的“巅峰”。从20世纪60 年代有关膳食碳水化合物(carbohydrate,CHO)与运动能力的突破性研究,到近些年有关细胞信号传导与肌肉在运动中的适应性反应的研究,结果表明,在训练或比赛的准备阶段控制膳食CHO 和脂肪可以有效地改变这些主要能源物质的可用性和利用率,从而提升运动表现[3]。早在2003 年,国际奥委会专家小组的官方建议就指出,不同类型的赛事或训练强度和训练量之间的能源物质需求不同,由此导致每日CHO 摄入量的目标会依具体情况而浮动,而不是一概建议始终摄入高CHO[4]。因此,2010 年更新的建议提出,应该使用CHO 可用性而不是绝对摄入量来评估运动员的膳食CHO 消耗量[5]。值得注意的是,除了早先被人们所熟知并广泛应用的高CHO 供应原则外,最新的运动营养指南强化了人们对低CHO 可用性的认识[1],进一步揭示了低CHO可用性训练的分子基础[2,6,7],并公布了在精英运动员中使用低CHO 可用性策略的案例研究[8],其中有研究证据表明,将低CHO可用性周密整合到训练有素运动员的训练计划中去,可以为他们带来卓越的运动表现[9,10]。事实上,最新的指导方针还指出通过周期化方法来控制训练饮食中的CHO 可用性可以带来众多的潜在益处[1,11]。
总体而言,营养与运动之间存在许多相互作用,从实践的角度来看,重要的是要了解在现实场景中如何适时地实施营养策略,以及在营养目标发生冲突时优先考虑何种重要策略,以优化特定的训练适应。因此,本文围绕CHO 周期化,进而延伸到脂肪,重点介绍通过控制CHO 可用性建立的新兴营养策略——“高训”(training with high-carbohydrate availability,高CHO可用性训练)策略,“低训”(training with low-carbohydrate availability,低CHO 可用性训练)策略,“低训高赛”(fat adaptation with carbohydrate restoration,脂肪适应与CHO 恢复相结合)策略,并分析关键的分子适应机制及对运动表现的影响,以期为未来如何最有效地将这种周期化营养策略纳入到运动训练计划的周期中去提供新的角度和思路。
1 周期化营养概述
1.1 来源及定义
周期化的概念深深植根于竞技运动的历史中,发展这一原则的开创性科学家和教练包括Hans Selye博士(提出“一般适应综合征”)[12],随后是Matveyev、Bonderchuck 和Bompa[13]。由此出现许多周期化的方法,包括传统模型、板块模型、极化模型和复杂模型[13,14]。这些多样性的方法中有一个核心主题:要有目的地合理安排不同的训练单元(大周期,以月计算;中周期,以周计算;短周期,以天计算),以便运动员能够达到所需的准备状态,在需要的目标赛事中表现最佳[15]。其中,训练适应理论是周期化训练的重要理论依据,而营养对训练适应和表现的影响至关重要。
基于此,2007 年国际田联营养共识首次正式提出了周期化营养的理论指导方针,提出了在年度阶段性训练计划中有必要按不同训练阶段的训练安排推荐能量摄入和宏量营养素需求[16]。运动训练中的周期化是一种长期渐进的方法,旨在通过全年系统地改变训练来提高运动成绩,而周期化营养通常用来描述在特定训练周期下营养摄入的变化[16-18]。例如,在某些训练期间,重点是体重管理和低能量摄入,而在其他训练期间,重点是加快恢复身体机能状态和优化运动表现以及更高的CHO 摄入。在周期化营养这一概念被逐渐认识的过程中,Hawley 和Burke 讨论了长期的“周期化训练—营养计划”作为一种提高运动成绩方式的重要性,指出竞技运动员应当在训练之前、期间或之后控制CHO 的可用性,这些是长期阶段性训练营养计划的一部分,旨在促进训练代谢适应,在理论上可以优化耐力运动表现[19]。Mujika等总结了营养应该定期进行调整,以支持在整个赛季和/或训练周期中不断变化的个人目标、训练水平和需求[18]。近些年,周期化营养被定义为有计划、有目的、战略性地使用特定的营养干预措施,以达到增强训练适应能力,或提升竞技表现等目的[11]。
1.2 CHO和脂肪周期化的常用方法
近些年,有关细胞信号传导的研究扩展了人们对营养支持在运动适应中促进作用的认识。许多底物,特别是肌糖原和血浆游离脂肪酸(free fatty acid,FFA),不仅可以作为运动的能源物质,还可以作为调节剂参与细胞和全身对运动的适应反应(尤其是耐力运动)。Hansen等[20]的开创性研究表明,对身体活动的一些适应可能需要肌糖原储备的“循环”,因此,CHO可用性在训练中的调节作用受到了越来越多的关注[8]。CHO 可用性周期化的概念可以用“为所需工作提供能源物质”的理论模型进行解释,其中CHO 可用性要根据所完成的特定训练任务/赛事的需求和目标进行调整[7]。通过这种方法,每日CHO 总摄入量及其在一天中的分布可以逐餐和逐日进行安排,以控制每次运动时的CHO 可用性。具体来说,“高CHO 可用性”(“高训”)表示这种摄入能够提供足够的内源性和/或外源性CHO供应以满足肌肉(和中枢神经系统)的需求,而“低CHO 可用性”(“低训”)表示供应和运动/能量需求之间的短缺[21]。此外,该领域的关注点还集中在“脂肪适应与高CHO恢复相结合”(“低训高赛”)这一周期化营养策略的功效上,涉及到在CHO 可用性降低的情况下完成特定的训练任务,以激活调节骨骼肌适应的分子途径,同时选择在CHO 可用性高的情况下进行关键训练任务和比赛,以实现运动表现和身体机能状态恢复的最大化[22,23]。然而,最重要的是要清楚CHO 周期化实际应用到整体运动训练计划中的最佳方法和时机(见表1,并详见下文)。

表1 CHO和脂肪周期化策略的常用方法
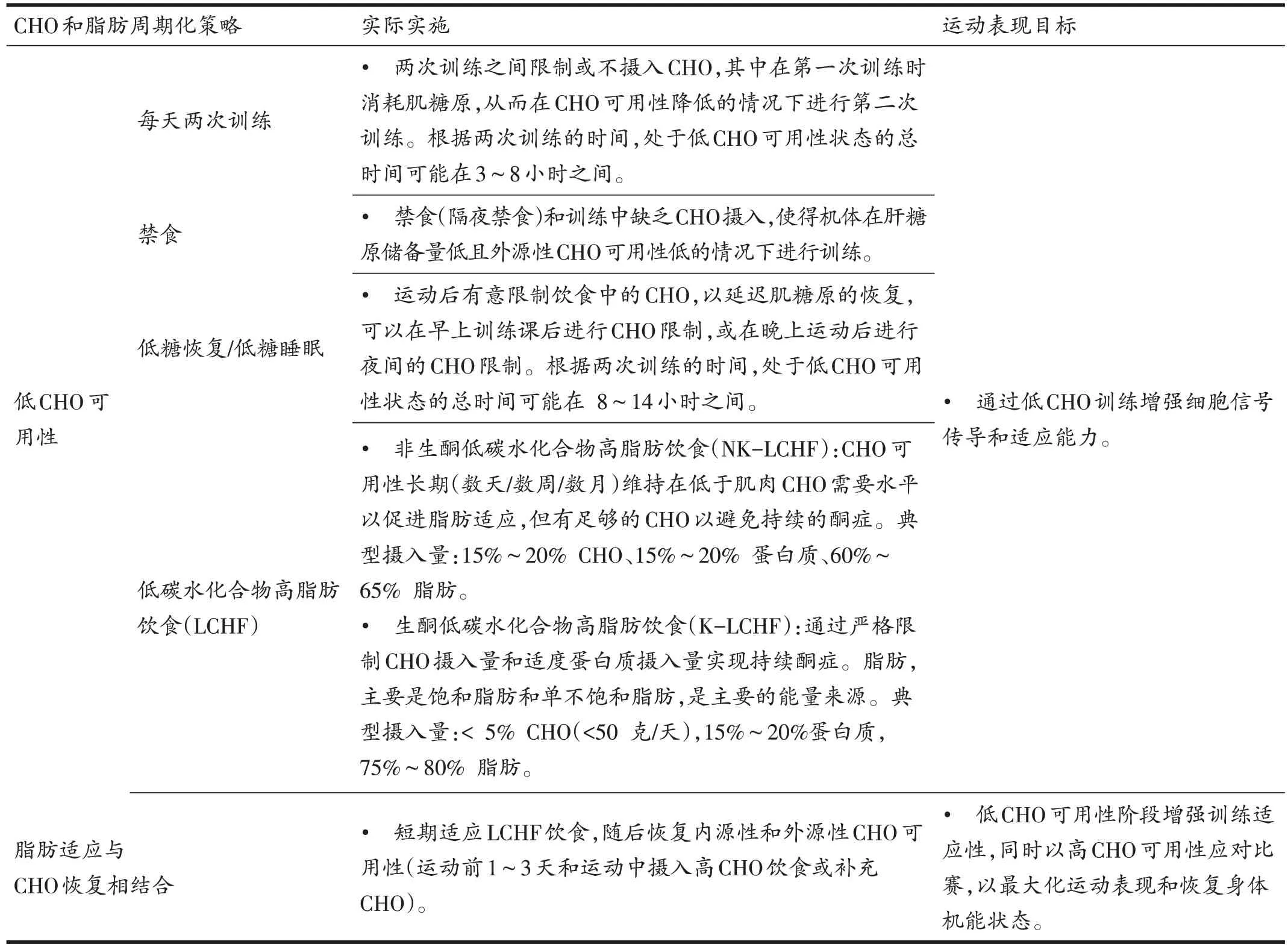
(续表1)
2 “高训”策略—高CHO可用性训练
“高训”策略是指在高CHO 可用性的条件下进行的训练,包括在运动开始时保持较高的肌糖原和肝糖原水平,和/或在运动期间补充CHO[11]。来自肌糖原储备和血浆葡萄糖的CHO 为各种运动表现提供了有效和主要的能量来源[24]。与100%脂肪氧化相比,每消耗1L 氧气,100% CHO 产生的三磷酸腺苷(adenosine-triphosphate,ATP)会多出5.5%[25]。CHO 作为肌肉和中枢神经系统的重要能源物质,其可用性限制了长时间亚极量或间歇性高强度运动的运动表现,并允许进行持续的高强度运动[26]。肌糖原的消耗与疲劳和持续运动强度的降低有关,而中枢神经系统的CHO 不足会对比赛成绩造成不利影响,如配速降低、运动技能和注意力受损以及运动中主观疲劳感知增加[27,28]。因此,实现高CHO 可用性,从而提供赛事或训练任务的能量需求是实施营养策略的一个关键目标[29-33]。
通过在训练前几小时或几天内补充CHO、在运动中摄入CHO 以及在训练间隙的恢复过程中补充能量,均可以增加CHO 可用性,这对于比赛或需要最佳表现的高强度训练非常重要[21]。受过耐力训练的个体中肌肉静息糖原的浓度高于久坐的个体,并且在受过训练的个体中可以实现糖原的超补偿(“CHO 负荷”),这涉及在24~48 小时内减少运动量以及提高CHO 的摄入(10~12 克/天/千克体重)[34]。这一策略一直被运动员用来提高其在耐力项目中的运动表现,以避免出现糖原耗竭的现象[3]。当然,许多运动对能源物质的需求超过了肌糖原的储存能力,或是说运动员在比赛间隙无法充分完成内源性储备的补充。基于此,提供额外的CHO是重要的,外源性CHO可以通过“节省”肝脏葡萄糖生成来维持正常血糖,随着糖原储备的耗尽,血糖对肌肉CHO 氧化速率的贡献不断增大[35]。有证据表明,在一系列运动场景中的运动表现可以通过在运动中摄入CHO来提高,摄入CHO的目的是补充肌肉所需的不断减少的糖原储备,且在这期间CHO 的摄入量可根据项目的特点进行调整:在长达2~3小时的耐力比赛项目中为30~60 克/小时;在持续8~10 小时的极限型耐力比赛项目中为60~90 克/小时[36];在持续1 小时的高强度运动中,少量的CHO,甚至包括使用含糖溶液漱口,都可以通过中枢神经系统效应提高运动能力[21]。此外,肠道对CHO的吸收可能是CHO氧化的限速步骤[37]。然而,在某种程度上,这可以通过“训练肠道”来克服:增加饮食和运动中CHO 的摄入,以增加耐受性和钠-葡萄糖共转运蛋白1(sodium-glucose cotransporter 1,SGLT-1)的活性[38,39]。使用可利用不同肠道转运蛋白的葡萄糖-果糖混合物也可以增加肠道对CHO的吸收和肌肉氧化率[40]。
3 “低训”(脂肪适应)策略——低CHO 可用性训练
“低训”策略是描述低CHO 可用性训练的一般术语,这种低CHO 可用性可以是低肌糖原/肝糖原,运动中或运动后的低CHO 摄入量,或两者的组合。控制糖原储备是优化训练适应性的一种有效手段。近些年,“低训”策略受到了越来越多的关注,包括一天两次训练策略、禁食策略、低糖恢复/低糖睡眠策略和低CHO高脂肪饮食。以下对不同方法的应用进行讨论,并通过分析低CHO 可用性调控训练适应的分子路径,分析其对骨骼肌代谢适应方面(如细胞信号传导、酶蛋白含量和活性的变化)的影响。
3.1 一天两次训练策略
“一天两次”训练策略是降低肌糖原储备的常用方案,包括在早晨的第一次训练时消耗肌糖原,然后在恢复过程中停止CHO 摄入,或者以较低水平摄入CHO,从而降低运动前CHO 的可用性,从而在低糖原状态下进行下午或傍晚的第二次训练。根据第一次和第二次训练之间的间隔时间(即训练间的恢复期)以及第二次训练的实际持续时间,累积的低糖原期可能在3~8小时之间。使用这一原理开展的第一项研究是Hansen等[20]的实验,他们通过使用单腿伸膝运动比较了每天一次的日常训练与每隔一天训练两次的效果,发现与始终处于CHO 负荷状态下的训练相比,一天两次训练显着改善了反映氧化能力的生物标志物[3-羟酰基辅酶A 脱氢酶(3-hydroxyacyl-CoA dehydrogenase,HAD)和柠檬酸合酶(citrate synthase,CS)]的活性,并提高了静息肌糖原水平。然而该模型并不能反映运动员现实训练/比赛中的情况。随后,由Hulston 等[41]和Yeo 等[42]进行的相同设计的平行研究观察了自行车运动员的干预情况。两项研究得出了相似的结果,即每隔一天训练两次的自行车运动员不能保持与每天训练一次的运动员相同的强度,尽管前者的做功较少,但训练适应能力却更强。此外,Morton等[43]也观察到在低肌糖原训练时发生的有益适应,琥珀酸脱氢酶(succinate dehydrogenase,SDH)活性增加。然而,与对照组相比,虽然3~10周的“低训”可提高氧化酶活性[20,41-43]、全身[41,42]和肌内脂肪利用率[41],但在五项涉及运动表现结果的研究中仅有两项显示出运动表现改善[20,44],其余的运动表现并无差异[41-43]。因此,一天两次的训练可能会引起机体发生有利于脂肪代谢的适应反应,然而这并不能转化为高强度运动期间运动表现的改善[43],且不足以证明这种训练方法会给长期的运动表现带来增益效果。
3.2 禁食策略
禁食策略代表了一种操作更为简单的“低训”模式,即在一夜禁食后,在空腹状态下进行训练。一般来说,最后一餐通常在前一天晚上的8点到10点之间,在第二天晨训过后再进食早餐。在该策略中,运动前肌糖原不会受到隔夜禁食的影响,但肝糖原会更低。相比之下,当运动强度、持续时间和完成的运动量一致时,FFA 可用性和脂质氧化会在禁食条件下出现增加[45]。实施禁食策略不仅可以增加急性运动后腺苷酸活化蛋白激酶[Adenosine 5’-monophosphate(AMP)-activated protein kinase,AMPK]的活性[46]和基因表达[47,48],几周的禁食训练还可以增加氧化酶活性[49]、脂质转运蛋白含量[50]、静息糖原储存[51],以及肌糖原再合成[52]。其中,Hespel 及其同事的研究表明[49,52],与进食状态下的训练(摄入含CHO 的早餐和在运动期间摄入CHO)相比,禁食状态下的训练能够引起机体产生更强的适应性。在6周训练(每周4次,75%VO2max下运动1~1.5小时)后,禁食状态下的氧化酶(如CS 和HAD)上调程度将进一步增加,其在提高肌肉氧化能力方面更为有效[49]。此外,他们还观察到肌内脂肪利用会随着禁食训练而出现增加,并注意到血糖水平的调节出现改善。以上研究结果表明在禁食状态下进行训练存在着潜在的益处。但需要注意的是,迄今为止关于禁食策略的研究主要集中在代谢适应上,很少有人报道其对运动表现带来的相关影响。随着时间的推移,禁食训练是否会改善运动表现还需要进一步观察。
3.3 低糖恢复/低糖睡眠策略
关于“低训”策略的另一个概念是在运动后的恢复期间限制CHO的摄入(“低糖恢复”)。Pilegaard等2005年进行了一项有趣的观察,结果显示,与对照组相比,当CHO 摄入受到限制超过5 小时时,75 分钟骑行运动后代谢基因的激活会出现增强[53]。然而,其他研究无法证实这一点,也没有发现恢复期间高CHO 摄入和低CHO摄入之间的差异[54,55]。随后出现的研究扩展了这一概念,通过晚上的高强度训练导致CHO(肌糖原和肝糖原)的可用性降低,然后在夜间睡眠期间继续限制CHO,因而又被称作“低糖睡眠”策略[59]。基于此,肌糖原减少的累计时间可延长至8~14 小时(这取决于训练时间和睡眠时间的长短)。这种做法违背了运动员在运动后(睡觉前)摄入CHO 以加速恢复的经典建议。然而“低糖睡眠”的急性模型(以糖原<200 mmol/kg 体重开始晨练)已被证实可以增强调节骨骼肌适应的分子途径[56-58],但当运动前后糖原含量分别为350和250 mmol/kg 体重时,这些反应会出现减弱[59]。Marquet 等[9,10]使用类似于Lane 等[59]的低糖睡眠模型观察到,与传统的“高训”策略相比,精英铁人三项运动员和自行车运动员进行1~3 周的低糖睡眠训练可以提高骑行效率(3.1%)、20公里骑行计时赛(time trail,TT)成绩(3.2%)和10 公里跑步成绩(2.9%)。目前,引发“低糖睡眠”最佳条件所需的CHO 限制程度还没有明确定义。此外,值得注意的是,这是仅有的两项关于“低糖睡眠”训练可以提升运动表现的研究[9,10],由此得出其可以优化运动表现的结论还为时过早。虽然这些研究提供了可以改善代谢适应的结果,但从实践的角度来看,在未来的研究中了解这种方法的其他潜在副作用(经常使用这一策略对身体机能恢复、免疫功能、睡眠质量和数量的影响)和未知因素十分重要。
3.4 非生酮/生酮低碳水化合物高脂肪饮食
与上述急性和间歇性/阶段性应用的“低训”策略不同,低碳水化合物高脂肪饮食(low-carbohydrate and high-fat diet,LCHF)是在限制CHO 的基础上增加了脂肪的摄入[60],并被建议为一种最大限度地将脂肪作为运动能源物质的慢性训练竞赛策略,包括非生酮(non-ketogenic,NK)-LCHF(通常60%~65%的能量来自脂肪,15%~20%的能量来自CHO,以防止酮症,同时将CHO 的摄入量减少到低于日常训练所需能源物质的水平)和限制更为严格的生酮(ketogenic,K)-LCHF(通常<50克/天CHO和75%~80%的脂肪)[3]。大量研究发现,LCHF 饮食能够上调肌肉中脂肪的释放、运输、摄取和利用,即使是在经过训练增强了适应能力的耐力运动员中也是如此[27,61]。值得注意的是,虽然肌肉脂肪代谢的最大变化发生在3~4 周内,但训练有素的运动员产生脂肪适应的时间过程似乎很快(5~10天)且稳定,生酮和非生酮的LCHF饮食都取得了相似的结果,由此将短期适应期定义为5~10天[61]。有强有力的证据表明,即使在精英运动员中,在坚持LCHF饮食数周/数月甚至数天内,肌肉的重组就可以提高运动中脂肪的使用(达到~1.5克/分钟),最大脂肪氧化强度能够从最大有氧能力的45% 变为70%[62]。此外,K-LCHF 饮食使肝脏产生更多酮体,为肌肉和中枢神经系统(central nervous system,CNS)提供了额外的能源物质,同时与血浆酮体(β-hydroxybutyrate,β-HB)的持续升高有关(β-HB >0.5 mmol/L)[63,64]。
然而,适应LCHF饮食并不会转化为耐力表现的明显改善,其益处大多是基于脂肪氧化的增加和循环酮体水平的上升[63,65]。有研究显示,自行车运动员适应这种CHO限制4周,增加了脂肪的利用,以维持次最大功率输出下的运动能力(~60% VO2max)[66]。然而,Phinney等注意到,这一结果是在高强度运动能力受损的情况下实现的。一项对精英竞走运动员进行的为期3.5周的K-LCHF 饮食调查验证了Phinney 等研究结果的合理性[67]。在这项研究中,最大脂肪氧化率是在适应K-LCHF 饮食后,在禁食或长时间运动期间摄入脂肪的条件下实现的。然而,这与在竞赛项目(20 和50 公里)中以一定速度行走的耗氧量增加(即经济性降低)有关,并且与应用CHO饮食组相比,K-LCHF饮食并没有提高10000米的成绩[67]。
目前,LCHF干预已经显示出潜在的局限性:(1)通过减少糖原利用和丙酮酸脱氢酶(pyruvate dehydrogenase,PDH)激活导致CHO 氧化受损[68-70];(2)脂肪氧化增加所带来的ATP 生成减少导致了运动经济性下降[67];(3)可能导致肠道SGLT-1减少,从而降低肠道吸收能力,这将损害CHO 补充策略的有效性,从而增加肠道紊乱的风险[38];(4)影响高强度运动的表现,即使是中等强度的运动,个体对LCHF 的反应也是不同的,提高/损害运动表现这两种结果的极端情况同时存在[62];(5)在接触K-LCHF饮食的最初几天,明显出现健康状况下降(例如疲劳、易怒、注意力不集中)和运动过程中主观疲劳感知增加(对于训练有素的运动员来说,训练能力通常在2~3周内才能恢复)[71]。因此,在实践中应用此类饮食干预,需要教练和运动员了解自身项目中的代谢需求和限制因素及其在个体水平上产生的特异性,从而在需要实现训练适应的阶段针对性地进行。
3.5 低CHO可用性调控训练适应的分子路径
在CHO 可用性降低的情况下有目的地进行特定训练任务的基本原理是,以低CHO 可用性状态开始部分训练和/或恢复时能够激活调节骨骼肌适应的细胞信号通路,该通路进一步促进机体产生与训练相关的适应性,如线粒体生物发生、血管生成和脂质氧化增加。糖原浓度是关键细胞信号激酶[例如,AMPK、P38丝裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)]、转录因子[例如,p53肿瘤抑制蛋白(p53 tumor suppressor,p53)、过氧化物酶体增殖物激活受体δ(peroxisome proliferator-activated receptor delta,PPARδ)]和转录辅激活因子[例如,过氧化物酶体增殖物激活受体γ辅激活子1α(peroxisome proliferator-activated receptor-γ coactivator 1α,PGC-1α)]的有效调节因素[6,58,72,73]。研究发现,在肌糖原减少的条件下进行运动可以增强AMPK 和p38MAPK 磷酸化,随后导致PGC-1α活化和转位到线粒体和细胞核中。然而,在运动前摄入富含CHO 的食物和/或在运动期间补充CHO可以下调脂质分解,并降低AMPK和p38MAPK活性,从而对下游调控因子产生负面影响[74](详见图1)。
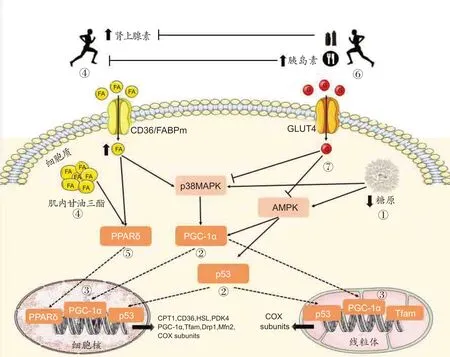
图1 低CHO可用性调控训练适应的分子路径
4 “低训高赛”策略—脂肪适应与CHO 恢复相结合
“低训高赛”策略试图“两全其美”地提供能源物质支持,通过短期适应LCHF 饮食以促进肌肉重组,从而增强FFA 运输和利用,随后恢复内源性和外源性CHO可用性(运动前1~3天和运动中摄入高CHO饮食或补充CHO)。该策略旨在将降低的CHO可用性周密整合到特定的训练任务中,以增强训练适应性,同时以高CHO 可用性应对比赛,以最大化运动表现和恢复身体机能状态[75,76]。有关研究表明,前期脂肪适应阶段实现的肌肉重组,即使在转为高CHO 可用性恢复后,依然能够促使机体持续地产生更高水平的脂肪氧化率[68,69,77]。尽管如此,没有研究发现随后的TT 成绩有明显提高,“低训高赛”策略对运动表现的益处仅适用于特定的情况或个体(表2)。此外,值得注意的是,由于糖原分解和PDH活性形式减少,CHO氧化的受损会阻碍运动前糖原储备的恢复[70]。Havemann等在实际竞赛场景中验证了这一观点:与进行高CHO 补充相比,自行车运动员在适应LCHF并恢复CHO可用性后,100公里骑行TT 成绩并未出现显着性变化。然而,当被要求以90%峰值功率输出(95%VO2peak)进行1 公里冲刺时,尽管主观疲劳感知和心率得以维持,但输出功率仍明显下降[69],这一结果可能表现为高强度运动下ATP 生成能力的降低。
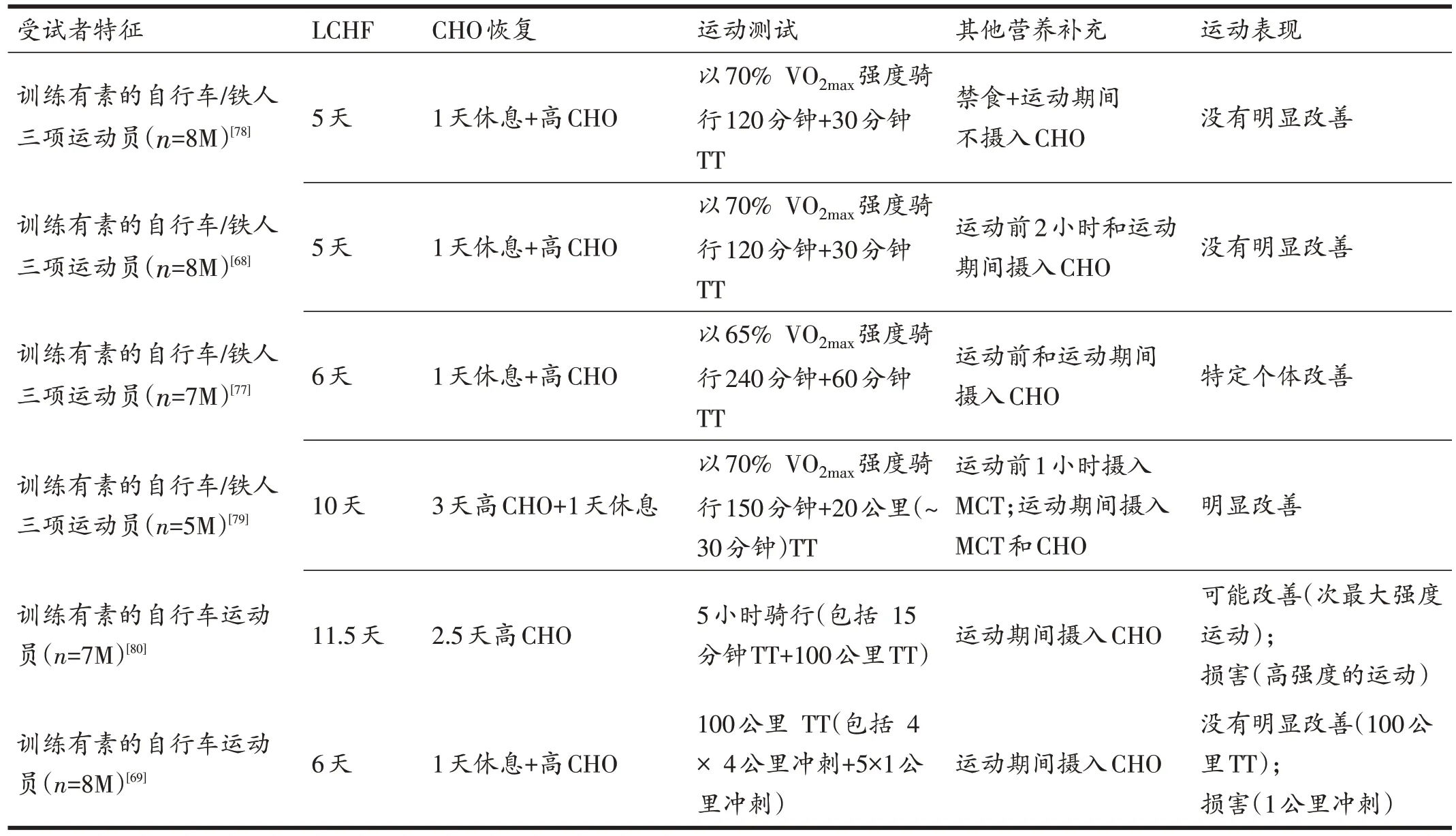
表2 “低训高赛”与运动表现的研究
5 结论与展望
在训练或比赛涉及更高强度的工作量/技战术以及需要最佳运动表现时,有必要使用“高训”策略提供高质量的输出和/或最大化的恢复;而在准备期训练或基于中低强度运动的训练负荷时,可能不需要高CHO可用性的支持。“低训”策略增强运动表现的科学证据尚不充分,但“低训高赛”策略对运动表现的益处可以适用于特定情况(长时间次最大强度运动)和特定个体(对CHO敏感的个体)。“低训高赛”策略会限制高强度运动的能力,但是配合低CHO 可用性有助于增强训练适应,在上调骨骼肌适应的分子路径上具有独特优势,通过这一过程实现的肌肉适应是优化运动表现的关键之一。将不同CHO 可用性的营养策略纳入到运动员的训练计划中有部分理论依据,但由于多种原因,平衡风险与回报可能具有挑战性。应该使用哪些策略取决于运动员个人的具体目标,没有任何一种策略(或饮食)可以满足运动员个人在所有情况下的所有需求,最佳的实践方法在于对不同的营养策略进行合理的周期化组合。因此,未来的核心问题在于如何以多元化的方式整合各类营养策略并充分加以利用,这就需要根据提高运动表现或是增强训练适应的优先级,分别考虑运动员在训练/比赛计划中使用高或低CHO 可用性的重要性,由此制定出一套个性化的营养策略,以实现运动员的个人整体营养目标,并最大限度地提高他们的训练和比赛成绩。



