任万平,邵 伟,2,雒诚龙,余 雄
(1.新疆农业大学动物科学学院,乌鲁木齐 830052; 2.新疆肉乳用草食动物营养重点实验室,乌鲁木齐 830052)
动物肌内脂肪的含量和积累使肌肉表现出大理石花纹状,影响和决定着肉的品质;尤其是高档雪花牛肉,肌肉中的脂肪含量和分布越丰富,牛肉的品质也就越好,因此,如何实现动物肌内脂肪的合成与沉积具有重要的现实和科学意义。磷酸二羟基丙酮(dihydroxyacetone phosphate,DHAP)作为糖脂代谢的重要中间产物,参与到生物代谢,其代谢途径主要有两条:(1)糖代谢生成的二羟基丙酮,经二羟基丙酮激酶磷酸化,生成磷酸二羟基丙酮,在磷酸丙糖异构酶的催化下,生成3-磷酸甘油醛,经一系列酶催化最终生成丙酮酸,进入三羧酸循环,再经过一系列酶促反应生成生脂原料乙酰辅酶A,用来合成脂肪;(2)二羟基丙酮经二羟基丙酮激酶磷酸化后,生成磷酸二羟基丙酮,经扩散作用进入细胞内,在α-磷酸甘油脱氢酶的作用下,还原成α-磷酸甘油,再经甘油转酰基酶催化,与脂酰辅酶A最终合成三酰甘油[1-2]。国内针对磷酸二羟基丙酮的研究主要是利用其合成稀有单糖[3-4],而磷酸二羟基丙酮直接使用对脂肪代谢的影响尚未见报道;作为其代谢前体的二羟基丙酮,用作新型饲料添加剂,用来提高生长降低体脂沉积和改善肉品品质[5-6]。
本试验旨在研究磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油及其关键代谢酶含量的影响,通过试验结果分析其对脂肪代谢的影响机制,为调控动物肌内脂肪的沉积提供理论依据。
1 材料与方法
1.1 试验材料
小鼠前脂细胞(3T3-L1)购于上海赛笠生物技术公司;磷酸二羟基丙酮(DHAP)、DMEM高糖培养基、胎牛血清(FBS)、蛋白消化液(EDTA)、磷酸盐缓冲液(PBS)、青链霉素试剂、3-异丁基-1-甲基黄嘌呤(IBMX)、地塞米松(DEX)、胰岛素(INS)购于美国GIBCO公司;试验所用ELISA试剂盒均由上海美轩生物技术公司生产。
1.2 试验设计
试验于2016年10月至2017年7月在新疆农业大学肉乳用草食动物营养重点实验室进行。按照单因素试验设计,在细胞培养液中分别加入不同剂量的磷酸二羟基丙酮作为不同试验组:20 μmol·L-1(试验Ⅰ组)、50 μmol·L-1(试验Ⅱ组)、200 μmol·L-1(试验Ⅲ组)以及不添加的空白对照组(ck组),并于0、36和72 h收集细胞及培养液冻存,以备送检;每组试验3个平行样,每个平行样3次重复。
1.3 测定指标及方法
1.3.1 测定指标 试验收集的细胞冻存后送由上海赛笠生物技术公司检测。检测项目:果糖-6-磷酸激酶(6-PFK)、醛缩酶(FDA)、葡萄糖-6-磷酸脱氢酶(G6PD)、丙酮酸脱氢酶E1(PDHE1)、二硫辛酸转乙酰基酶(DLAT)、二硫辛酰胺脱氢酶(DLD)、乙酰CoA羧化酶(ACC)、脂肪酸合成酶(FAS)、甘油二酯转移酶2(DGAT2)、激素敏感脂肪酶(HSL)、三酰甘油水解酶(TGH)、肉碱脂酰转移酶Ⅰ(CPT1)、三酰甘油(TG)、甘油二酯(DG)、游离脂肪酸(FFA)、丙酮酸(PA)、柠檬酸(CA)。
1.3.2 试剂配制 完全培养基:89%DMEM高糖培养基+10%FBS胎牛血清+1%青链霉液;诱导液Ⅰ:完全培养基+0.5 mmol·L-1IBMX+1 μmol·L-1DEX+5 mg·L-1INS;诱导液Ⅱ:完全培养基+5 mg·L-1INS。
1.3.3 细胞培养 参照郭秀玲等[7]方法将小鼠前脂细胞(3T3-L1)于含10%胎牛血清的完全培养基中,在37 ℃、5%CO2饱和湿度条件下培养,待细胞融合度达80%左右,将细胞传代接种到培养板中常规培养,融合度达80%时诱导,先用诱导液Ⅰ培养2 d,再换诱导液Ⅱ培养2 d,更换完全培养基培养第14天,加入含0、20、50、200 μmol·L-1的磷酸二羟基丙酮完全培养基培养细胞,于0、36、72 h收集细胞和培养基进行后续检测。
1.3.4 代谢产物和酶含量的测定 试验中各种代谢产物和酶含量的测定均参照闫莉等[8]的双抗体夹心法。每个样品平行测定3次。
1.4 统计方法
数据用“平均值(Means)±标准差(SD)”表示,用SPSS22进行单因素方差分析,并用LSD方法进行多重比较。
2 结 果
2.1 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油合成中关键限速酶含量的影响
由表1 可知,在36和72 h时,试验Ⅰ、Ⅱ、Ⅲ组的G6PD水平均极显着高于对照组(P<0.01),且试验各组之间差异不显着(P>0.05);在36和72 h时,试验各组DLAT、DLD和FAS的含量均极显着高于对照组(P<0.01),试验各组之间差异不显着(P>0.05);试验各组ACC含量差异均不显着(P>0.05);在36和72 h时,试验各组DGAT2的含量显着或极显着高于对照组(P<0.05,P<0.01),试验各组之间差异不显着(P>0.05)。
表1磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油合成中关键限速酶含量的影响
Table1Effectofdihydroxyacetonephosphateonkeyrate-limitingenzymecontentsinTGsynthesisinmouseadipocytes
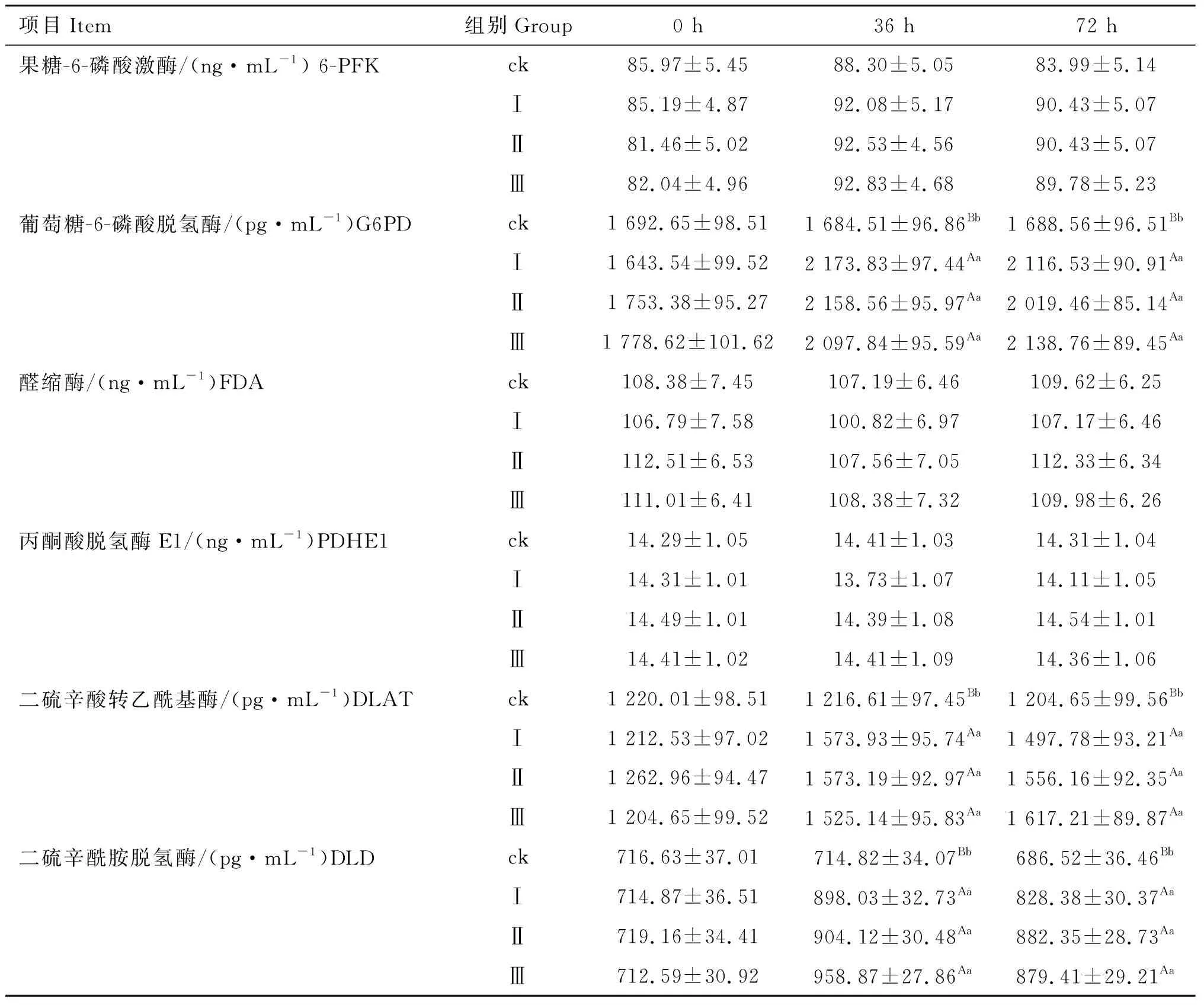
项目Item组别Group0 h36 h72 h果糖-6-磷酸激酶/(ng·mL-1) 6-PFKck85.97±5.4588.30±5.0583.99±5.14Ⅰ85.19±4.8792.08±5.1790.43±5.07Ⅱ81.46±5.0292.53±4.5690.43±5.07Ⅲ82.04±4.9692.83±4.6889.78±5.23葡萄糖-6-磷酸脱氢酶/(pg·mL-1)G6PDck1 692.65±98.511 684.51±96.86Bb1 688.56±96.51BbⅠ1 643.54±99.522 173.83±97.44Aa2 116.53±90.91AaⅡ1 753.38±95.272 158.56±95.97Aa2 019.46±85.14AaⅢ1 778.62±101.622 097.84±95.59Aa2 138.76±89.45Aa醛缩酶/(ng·mL-1)FDAck108.38±7.45107.19±6.46109.62±6.25Ⅰ106.79±7.58100.82±6.97107.17±6.46Ⅱ112.51±6.53107.56±7.05112.33±6.34Ⅲ111.01±6.41108.38±7.32109.98±6.26丙酮酸脱氢酶E1/(ng·mL-1)PDHE1ck14.29±1.0514.41±1.0314.31±1.04Ⅰ14.31±1.0113.73±1.0714.11±1.05Ⅱ14.49±1.0114.39±1.0814.54±1.01Ⅲ14.41±1.0214.41±1.0914.36±1.06二硫辛酸转乙酰基酶/(pg·mL-1)DLATck1 220.01±98.511 216.61±97.45Bb1 204.65±99.56BbⅠ1 212.53±97.021 573.93±95.74Aa1 497.78±93.21AaⅡ1 262.96±94.471 573.19±92.97Aa1 556.16±92.35AaⅢ1 204.65±99.521 525.14±95.83Aa1 617.21±89.87Aa二硫辛酰胺脱氢酶/(pg·mL-1)DLDck716.63±37.01714.82±34.07Bb686.52±36.46BbⅠ714.87±36.51898.03±32.73Aa828.38±30.37AaⅡ719.16±34.41904.12±30.48Aa882.35±28.73AaⅢ712.59±30.92958.87±27.86Aa879.41±29.21Aa
(转下页 Carried forward)
各试验组间,同列数据肩标不同大写字母表示差异极显着(P<0.01),肩标不同小写字母表示差异显着(P<0.05),字母相同或无肩标表示差异不显着(P>0.05)。下同
In the same column,values with different capital letter superscripts mean extremely significant difference(P<0.01), values with different small letter superscripts mean significant difference(P<0.05), while values with the same or no letter superscript mean no significant difference(P>0.05) among different groups. The same as below
2.2 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油分解代谢中关键限速酶含量的影响
由表2可知,在36和72 h时,试验Ⅰ、Ⅱ、Ⅲ组HSL含量均极显着低于对照组(P<0.01),试验各组之间差异不显着(P>0.05);试验各组TGH含量与对照组差异均不显着(P>0.05);在36和72 h时,试验各组CPT1含量均极显着低于对照组(P<0.01),试验各组之间差异不显着(P>0.05)。
表2磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油分解代谢中关键限速酶含量的影响
Table2Effectofdihydroxyacetonephosphateonkeyrate-limitingenzymecontentsinTGcatabolisminmouseadipocytes
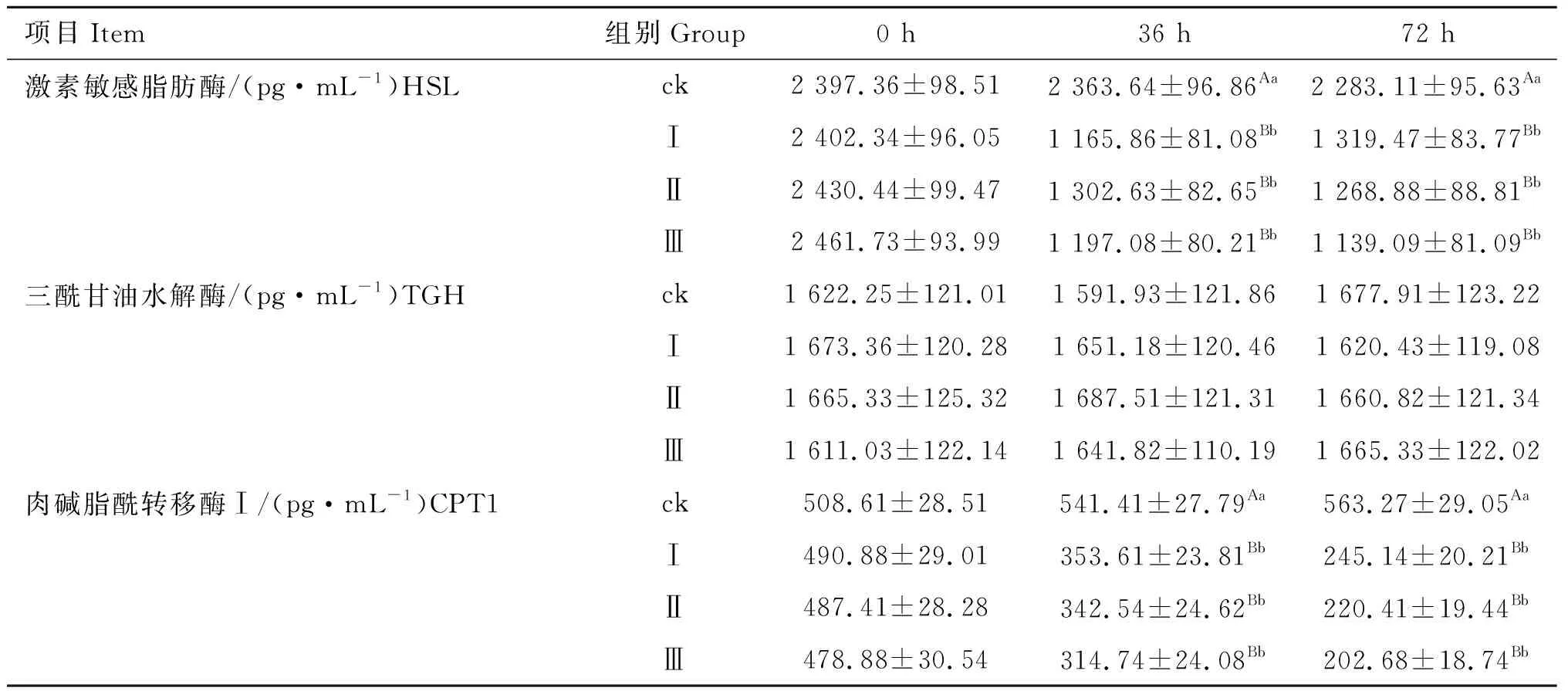
项目Item组别Group0 h36 h72 h激素敏感脂肪酶/(pg·mL-1)HSLck2 397.36±98.512 363.64±96.86Aa2 283.11±95.63AaⅠ2 402.34±96.051 165.86±81.08Bb1 319.47±83.77BbⅡ2 430.44±99.471 302.63±82.65Bb1 268.88±88.81BbⅢ2 461.73±93.991 197.08±80.21Bb1 139.09±81.09Bb三酰甘油水解酶/(pg·mL-1)TGHck1 622.25±121.011 591.93±121.861 677.91±123.22Ⅰ1 673.36±120.281 651.18±120.461 620.43±119.08Ⅱ1 665.33±125.321 687.51±121.311 660.82±121.34Ⅲ1 611.03±122.141 641.82±110.191 665.33±122.02肉碱脂酰转移酶Ⅰ/(pg·mL-1)CPT1ck508.61±28.51541.41±27.79Aa563.27±29.05AaⅠ490.88±29.01353.61±23.81Bb245.14±20.21BbⅡ487.41±28.28342.54±24.62Bb220.41±19.44BbⅢ478.88±30.54314.74±24.08Bb202.68±18.74Bb
2.3 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油代谢关键产物的影响
由表3可知,在36和72 h时,试验各组TG含量均极显着高于对照组(P<0.01),且试验各组之间差异不显着(P>0.05);在36和72 h时,试验各组DG含量均显着低于对照组(P<0.05),试验各组之间差异不显着(P>0.05);在36和72 h时,试验各组FFA含量均极显着低于对照组(P<0.01),试验各组之间差异不显着(P>0.05);在36 h时,试验各组PA含量与对照组差异不显着(P>0.05),在72 h 时,试验各组PA含量均显着高于对照组(P<0.05);在36和72 h时,试验各组CA含量均极显着高于对照组(P<0.01),试验各组之间差异不显着(P>0.05)。
表3磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油代谢产物含量的影响
Table3EffectofdihydroxyacetonephosphateonthecontentsofTGmetabolitesinmouseadipocytes
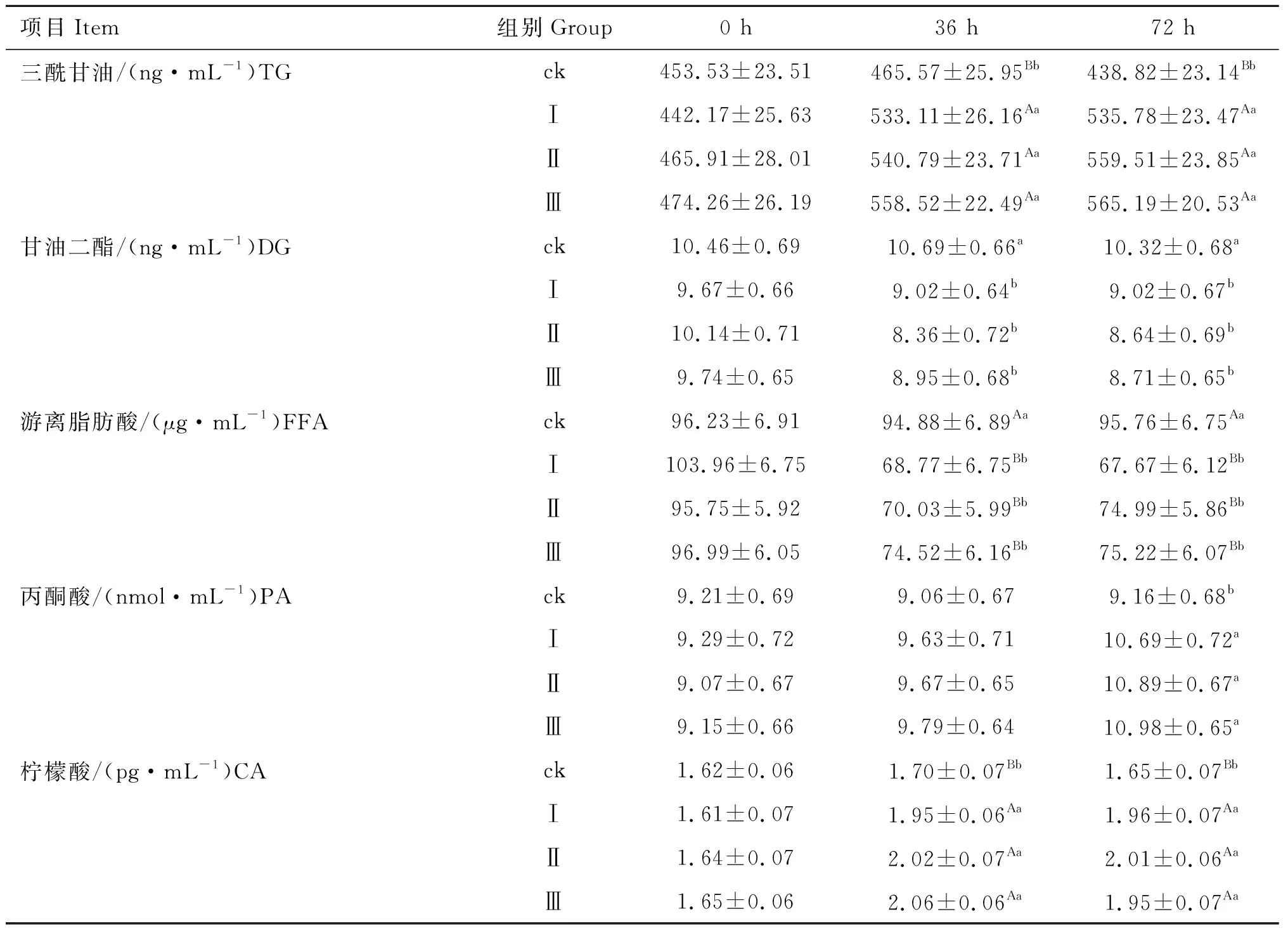
项目Item组别Group0 h36 h72 h三酰甘油/(ng·mL-1)TGck453.53±23.51465.57±25.95Bb438.82±23.14BbⅠ442.17±25.63533.11±26.16Aa535.78±23.47AaⅡ465.91±28.01540.79±23.71Aa559.51±23.85AaⅢ474.26±26.19558.52±22.49Aa565.19±20.53Aa甘油二酯/(ng·mL-1)DGck10.46±0.6910.69±0.66a10.32±0.68aⅠ9.67±0.669.02±0.64b9.02±0.67bⅡ10.14±0.718.36±0.72b8.64±0.69bⅢ9.74±0.658.95±0.68b8.71±0.65b游离脂肪酸/(μg·mL-1)FFAck96.23±6.9194.88±6.89Aa95.76±6.75AaⅠ103.96±6.7568.77±6.75Bb67.67±6.12BbⅡ95.75±5.9270.03±5.99Bb74.99±5.86BbⅢ96.99±6.0574.52±6.16Bb75.22±6.07Bb丙酮酸/(nmol·mL-1)PAck9.21±0.699.06±0.679.16±0.68bⅠ9.29±0.729.63±0.7110.69±0.72aⅡ9.07±0.679.67±0.6510.89±0.67aⅢ9.15±0.669.79±0.6410.98±0.65a柠檬酸/(pg·mL-1)CAck1.62±0.061.70±0.07Bb1.65±0.07BbⅠ1.61±0.071.95±0.06Aa1.96±0.07AaⅡ1.64±0.072.02±0.07Aa2.01±0.06AaⅢ1.65±0.062.06±0.06Aa1.95±0.07Aa
3 讨 论
3.1 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油合成关键限速酶含量的影响
三酰甘油的存在是细胞将糖转化为脂肪储存起来的结果,其实质就是糖脂代谢过程中一系列的酶促反应,葡萄糖经过一系列关键限速酶的催化生成乙酰辅酶A,生成的乙酰辅酶A再经过乙酰辅酶A羧化酶、脂肪酸合成酶催化生成脂肪酸,再与糖代谢生成的中间产物α-磷酸甘油经过甘油酰基转移酶系的催化,最终生成三酰甘油,因此任何糖、脂代谢中关键限速酶含量与中间产物含量的变化都能直接影响到三酰甘油的代谢。G6PD是糖脂代谢过程中的关键限速酶,能够催化葡萄糖生成还原性辅酶A(NADPH)、乙酰辅酶A等用于脂肪酸的合成。本研究中(表1),在36和72 h时,试验各组的G6PD均极显着高于对照组(P<0.01),郑雅楠等[9]研究发现,G6PD表达增高,能够促进NADPH的含量增加,而Khu等[10]研究发现,糖代谢中G6PD升高,能够促进细胞对糖的利用,增加脂肪的合成和沉积,这与本研究结果一致。PDHE1、DLAT、DLD统称丙酮酸脱氢酶复合体,由葡萄糖代谢产生的丙酮酸经此酶系催化脱羧生成乙酰CoA,DLAT和DLD在该酶系中起到主要调节作用。在36和72 h时,试验各组DLAT和DLD的含量极显着高于对照组(P<0.01),程钰蓉等[11]研究发现,丙酮酸脱氢酶系活性增加,脂肪合成增强,其活性抑制能够降低脂肪的合成,与本研究结果相似,说明添加不同剂量的磷酸二羟基丙酮能够提高细胞中丙酮酸脱氢酶系的活性,从而促进丙酮酸的代谢。
以上糖代谢的酶促反应中,由G6PD催化反应产生的NADPH+H+为脂肪的生物合成提供了所需的大量氢体,丙酮酸脱氢酶复合体则催化丙酮酸生成脂肪合成的原料乙酰CoA,这些由糖代谢生成的氢体和乙酰CoA存在于胞液中,接下来再经过脂代谢关键合成酶ACC与FAS的催化,生成脂肪酸链,用于合成三酰甘油。本研究中,试验各组的G6PD与丙酮酸脱氢酶复合体的含量极显着提高,说明试验各组生成的生脂原料和氢体增多。
ACC能够催化乙酰CoA生成丙二酰辅酶A,是催化合成脂肪酸的第一步,也是最重要的一步,生成的丙二酰辅酶A的含量直接决定着脂肪酸的合成量。本研究中(表1),添加磷酸二羟基丙酮的试验各组能够提高ACC的含量,在36和72 h时,试验Ⅰ组ACC的含量就比对照组分别高出了9.62%和9.69%,李亮等[12]研究发现,ACC的含量升高,机体内脂肪的合成沉积增加;Mao等[13]和Barber等[14]研究证实,ACC活性增加能够促进细胞三酰甘油的合成与沉积,这与本研究结果一致。FAS是一个多酶复合体,催化乙酰CoA生成脂肪酸。试验结果表明,添加不同剂量磷酸二羟基丙酮能够极显着提高FAS的含量(P<0.01),刘作华等[15]研究发现,FAS表达升高,脂肪的含量增加,而Dentin等[16]研究也证实其活性的高低直接控制脂肪合成的强弱,其表达水平升高能够显着增加三酰甘油的沉积,充分证明了本研究结果。在ACC和FAS催化的脂代谢反应中,本研究中试验各组的ACC和FAS极显着高于对照组,说明试验各组合成的脂肪酸量也显着高于对照组。DGAT2是催化甘油二酯转变成三酰甘油的限速酶,能够催化α-磷酸甘油(由磷酸二羟基丙酮还原得到)和脂肪酸生成三酰甘油。由本试验结果可知,在36 h时,试验各组DGAT2含量显着高于对照组(P<0.05),72 h时,极显着高于对照组水平(P<0.01);Nestel[17]、张晓图等[18]研究发现,DGAT2表达和活性升高,三酰甘油合成加强,这与本研究结果一致。因为试验各组的三酰甘油合成量强于对照组,所以试验各组的三酰甘油含量也显着高于对照组。
3.2 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油分解代谢中关键限速酶含量的影响
三酰甘油的存在是一个合成和分解动态平衡的结果,其在合成的同时分解也在进行。HSL在脂肪分解中起到决定性作用,是分解脂肪的关键限速酶。本研究中(表2),试验各组HSL的含量均极显着低于对照组水平(P<0.01),这与Lorente-Cebrin等[19]与Xu等[20]研究结果一致,HSL表达抑制,脂肪分解减少;说明磷酸二羟基丙酮能够极显着降低HSL含量和活性,从而抑制三酰甘油的分解。TGH也是催化三酰甘油水解的重要脂肪酶,Cornaciu等[21]、Chakrabarti和Kandror[22]和Serr等[23]研究发现,TGH的表达抑制能够促进三酰甘油的沉积,何微[24]研究发现,高脂营养水平促进脂肪合成和沉积时,TGH表达显着被抑制,而袁禹惠[25]研究发现,高脂营养水平能够抑制脂肪的沉积,但是对TGH酶的表达和含量影响并不显着,这与本研究结果一致,表明添加磷酸二羟基丙酮对TGH整体含量影响不显着(P>0.05)。
肉碱脂酰转移酶Ⅰ(CPT1)是转运活化的脂肪酸(脂酰CoA)进入线粒体进行氧化分解的关键限速酶。本研究中(表2),试验各组CPT1的含量在36和72 h时均极显着低于对照组(P<0.01),试验各组之间差异均不显着(P>0.05),胡伟[26]、董婧[27]、Abu-Elheiga等[28]研究证实,抑制CPT1的表达和含量能够抑制脂肪的氧化,增加脂肪的沉积,与本研究结果一致,说明磷酸二羟基丙酮能够降低脂肪细胞内CPT1的含量,抑制脂肪酸的β氧化分解,从而增加三酰甘油在脂肪细胞内的沉积。
本研究中,试验各组HSL和CPT1含量均极显着低于对照组,说明试验各组生成的三酰甘油分解速率也显着低于对照组,这也是试验各组三酰甘油含量显着高于对照组的原因之一。
3.3 磷酸二羟基丙酮对小鼠脂肪细胞三酰甘油代谢关键产物含量的影响
本研究结果可知(表3),试验各组TG的含量在36和72 h时均极显着高于对照组(P<0.01),试验各组之间差异不显着(P>0.05),推测可能是磷酸二羟基丙酮一方面通过提高ACC、FAS酶的活性,促进FFA的合成;另一方面磷酸二羟基丙酮经过还原作用生成α-磷酸甘油,与之前生成的FFA经甘油二酯转移酶系催化生成三酰甘油,而本研究中FFA含量极显着降低,DGAT2酶含量极显着升高能够说明这些。杨少鹏[2]证实,磷酸二羟基丙酮在生物体内一是参与糖代谢,二是用于合成三酰甘油[29],也能够证明本研究结果,说明磷酸二羟基丙酮能够促进小鼠脂肪细胞三酰甘油的合成和沉积。
DG和FFA既是TG的合成前体同时也是其分解产物,在本研究中,试验各组FFA的含量均极显着低于对照组(P<0.01),试验各组DG含量均显着低于对照组(P<0.05),这与杨竹青等[30]通过烟酸抑制三酰甘油分解从而显着降低细胞中FFA含量结果一致,因为添加磷酸二羟基丙酮能够显着降低三酰甘油水解酶HSL的含量,极显着提高DGAT2和TG含量,更多的FFA和DG用于合成TG,因此试验各组FFA和DG的含量显着降低。
PA、CA作为糖脂代谢的关键中间产物,构成柠檬酸-丙酮酸循环,能够将合脂原料乙酰CoA从线粒体中转运至胞液中,用于合成脂肪。在本研究中,试验各组PA的含量在72 h均高出对照组水平(P<0.05),试验各组CA含量在36和72 h均极显着高出对照组水平(P<0.01),推测,一是添加的磷酸二羟基丙酮本身参与代谢,生成更多PA和CA,二是由于添加了磷酸二羟基丙酮,多余的葡萄糖用于代谢生成PA和CA,因此能够生成并转运更多的生脂原料—乙酰CoA,Wang等[31]研究发现,在脂代谢的调控过程中,多余的葡萄糖会诱导PA、CA、ACC及FAS等表达,从而促进脂肪的合成,这与本研究结果一致。
4 结 论
外源性添加磷酸二羟基丙酮能够极显着提高小鼠脂肪细胞三酰甘油合成代谢中G6PD、DLAT、DLD、FAS等酶的含量,同时极显着降低三酰甘油分解代谢中HSL与CPT1酶的含量;从而增强小鼠脂肪细胞中三酰甘油的含量和沉积。



