李 琛,何文峰,赵丽娜,凡 启,杨国庆,刘慧敏
(河南农业大学生命科学学院,郑州 450002)
伪狂犬病(pseudorabies,PR)是由伪狂犬病病毒(pseudorabies virus,PRV)引起的可感染多种家畜和哺乳动物的急性传染病,以发热、奇痒(猪除外)及急性脑脊髓炎为主要症状。PRV属于疱疹病毒科α-疱疹病毒亚科。猪是PRV的唯一自然感染宿主以及长期贮存和传播宿主。PRV感染猪后可建立持续性感染,主要潜伏在猪的外周神经组织,在机体免疫低下或受到应激时可被激活,导致机体发病甚至成为传染源。PRV特殊的致病方式和感染方式给该病的防控带来较大困难,严重危害全球养猪业的发展。近年来,PRV更是突破了常规疫苗,导致猪伪狂犬病的防控难度极大,成为养猪业防控伪狂犬病所面临的巨大挑战。
在病毒入侵机体过程中,宿主产生各种防御机制来保护机体,先天性免疫应答是抵抗入侵病原体的第一道防线。PRV对宿主细胞的感染力很强,能够引起宿主细胞产生免疫反应,诱导产生Ⅰ型IFN,分泌的IFN与其受体结合激活下游信号转导,最终导致数百个干扰素刺激基因(ISGs)的表达。其中,ISG15(interferon-stimulated gene15)作为诱导最强烈、反应最快的ISG蛋白之一,在宿主抵御病毒感染的过程中发挥重要的作用。
近年来,对ISG15抗病毒复制的作用有越来越多的研究。ISG15可以通过作用于病毒入侵的各阶段影响病毒的增殖,也可以作为免疫调节蛋白来调控宿主的反应。本实验室前期研究发现PRV感染能显着上调ISG15,过表达ISG15能明显抑制PRV复制。但是ISG15抑制PRV复制的分子机制尚不清楚,有待进一步研究。在前期试验基础上,本研究利用CRISPR/Cas9基因编辑技术,构建15基因敲除PK-15细胞系(PK15-ISG15),利用PRV毒株检测敲除15基因对PRV复制的影响,以期为进一步研究ISG15抑制PRV 复制的分子机制提供新思路,同时为有效防控猪伪狂犬病提供新的研究策略。
1 材料与方法
1.1 试验材料
猪肾上皮细胞(PK-15)、PRV、ISG15兔多克隆抗体由作者实验室保存;pCompass-LVKO 载体购自Addgene;载体质粒pMD2.G和psPAX2购自Sigma;普通琼脂糖凝胶DNA回收试剂盒、质粒小提试剂盒均购自天根生化科技有限公司;限制性内切酶Ⅰ、RNAiso Plus、PrimeScriptRT reagent Kit with gDNA Eraser、Primer STARMax DNA Polymerase购自宝日医生物技术有限公司;MonAmpSYBRGreen qPCR Mix试剂购自莫纳生物科技有限公司;PRV鼠源gE单克隆抗体购自普莱柯生物工程股份有限公司;β-actin抗体购自赛维尔生物科技有限公司;Cell Counting Kit-8购自生工生物工程股份有限公司。
1.2 引物设计与合成
根据NCBI网站所公布的猪15基因序列(GenBank No.:NM_001128469),在15的单一外显子上找到2个合适的靶位点,将此靶序列与猪的基因组序列进行比对,仅在15基因座上找到完全匹配序列,针对2个位点分别设计sgRNA上游引物与下游引物,同时,在基因组靶序列两侧设计15的验证引物(表1);通过NCBI数据库查询需要检测的基因 mRNA序列,应用Primer-Blast设计RT-qPCR引物(表1)。

表1 引物序列及相关信息Table 1 Sequences and information of primers used in our study
1.3 ISG15基因敲除细胞系的构建
1.3.1 敲除载体的构建 将两对sgRNA引物95 ℃ 5 min进行退火杂交,退火后连接至Ⅰ限制性内切酶线性化的pCompass-LVKO载体中。重组载体送北京擎科生物科技有限公司测序,将测序正确质粒命名为pCompass-LVKO-ISG15-1、pCompass-LVKO-ISG15-2,转化至大肠杆菌DH5α感受态细胞中,挑取单菌落扩大培养,提取质粒后,-20 ℃保存备用。
1.3.2 敲除细胞系的构建 将pCompass-LVKO-ISG15-1、pCompass-LVKO-ISG15-2与psPAX2、pMD2G以2∶1∶1的比例,共转染HEK293T细胞;培养48 h后收集包装病毒上清液,取1 mL感染PK-15细胞,感染48 h后,用2 μg·mL嘌呤霉素筛选细胞,使用胰酶消化存活细胞,梯度稀释后,按照每孔1个细胞的比例接种至96孔板,37 ℃,5% CO培养扩增细胞。取15完全敲除的克隆,抽提基因组DNA后以此为模板,使用验证引物ISG15 VerF和ISG15 VerR进行PCR扩增,20 μL反应体系:PrimerSTAR Max Premix 10 μL,上、下游引物(0.2 μmol·L)各0.4 μL,模板1 μL,ddHO 8.2 μL。反应条件:98 ℃ 2 min;98 ℃ 10 s,55 ℃ 5 s,72 ℃ 5 s,共35个循环;72 ℃ 5 min。如果在619 bp处出现一条特异性条带,可初步说明敲除15细胞构建成功,将此条带连接至pEASY-T1 simple载体,随机挑选阳性克隆送公司测序。
1.4 ISG15敲除效率的验证
6孔板铺种PK-15和PK15-ISG15细胞,各设3组重复。培养至细胞汇合度70%左右接种病毒,以PRV MOI=2感染细胞,感染24 h收取蛋白,BCA法检测蛋白浓度。取35 μg蛋白样品进行SDS-PAGE电泳后转膜,5%脱脂奶室温封闭1 h后,加入ISG15兔多克隆抗体(1∶5 000)4 ℃摇床孵育过夜,TBST洗膜3次,室温孵育HRP标记的羊抗兔IgG(1∶5 000)1 h,ECL显影。以β-actin作为内参。
1.5 细胞增殖(CCK-8)检测
以每孔1×10个细胞在96孔板中铺种PK-15和PK15-ISG15细胞,每个处理设置5个复孔,培养24 h。参照CCK-8试剂盒在不同时间点向指定培养孔中加入10 μL的CCK-8溶液,37 ℃孵育2 h,随后用酶标仪检测在450 nm处的吸光度。试验重复3次。
1.6 间接免疫荧光检测
12孔板铺种PK-15和PK15-ISG15细胞,各3个重复,培养至细胞汇合度70%左右进行病毒处理,以PRV MOI=2感染细胞,24 h后取出,PBS洗涤两次,然后使用4%多聚甲醛室温固定15 min,使用破膜液渗透15 min。然后用封闭缓冲液(含5%脱脂奶粉的PBS)封闭细胞1 h后,加入鼠源PRV-gE抗体(1∶1 000)4 ℃孵育过夜。PBS清洗后,室温避光孵育FITC标记的山羊抗小鼠抗体(1∶200),PBS清洗3遍。滴入封片剂平铺覆盖细胞,倒置荧光显微镜下观察。
1.7 实时荧光定量PCR检测
6孔板铺种PK-15和PK15-ISG15细胞,各3组重复。培养至细胞汇合度70%左右进行病毒处理,以PRV MOI=2感染细胞,分别在0、6、12、24、36和48 h收取RNA。TRizol法提取RNA并反转录成cDNA,以cDNA为模板,进行RT-qPCR检测。每个样品3个重复。反应体系20 μL:MonAmpSYBR Green qPCR Mix 10 μL,上、下游引物(0.2 μmol·L)各0.4 μL,cDNA 3 μL,ddHO 6.2 μL。反应条件:95 ℃预变性10 min;95 ℃变性10 s,60 ℃退火和延伸30 s,共40个循环。以-mRNA表达水平为内参值,计算PRV-0、PRV-、PRV-16、-β mRNA水平。
1.8 Western blot检测PRV-gE蛋白的表达
6孔板铺种PK-15和PK15-ISG15细胞,各3组重复。培养至细胞汇合度70%左右进行病毒处理,以PRV MOI=2感染细胞,感染0、6、12、24、36和48 h收取蛋白,BCA法测蛋白浓度。取35 μg蛋白样品进行SDS-PAGE电泳后转膜,5%脱脂奶室温封闭1 h后,加入ISG15兔多克隆抗体(1∶5 000)4 ℃摇床孵育过夜,TBST洗膜3次,室温孵育HRP标记的羊抗兔IgG(1∶5 000)1 h后ECL显影。以β-actin作为内参。Image J软件分析条带灰度值比较ISG15敲除对PRV-gE蛋白表达的影响。
1.9 PRV子代病毒滴定测定
取对数生长期PK-15和PK15-ISG15细胞接种于6孔板。培养至细胞汇合度70%左右,弃去细胞培养物,PBS洗2遍,将稀释好的PRV病毒液400 μL接种到各孔中,置于37 ℃、5% CO培养箱1 h,期间每隔15 min摇晃一次,以便病毒吸附;弃去病毒液,加入含1%琼脂糖细胞维持液覆盖各孔,置于37 ℃、5% CO培养箱培养3 d,取出细胞培养板,10%福尔马林固定30 min,弃去细胞覆盖物,PBS洗2遍,用0.5%结晶紫染色30 min,选择合适的稀释度进行计数。
1.10 ISG15与PRV共孵育试验
取对数生长期PK-15细胞稀释接种于6孔板。培养至细胞融合度70%左右,进行分组处理:第1组直接用PRV(MOI=2)感染细胞;第2组将PRV与BSA于37 ℃孵育1 h后加入细胞;第3组将PRV与ISG15蛋白孵育1 h后加入细胞。吸附1 h后,更换成2%维持培养基,24 h后收取细胞总RNA、总蛋白和上清。分别检测PRV-gE的mRNA和蛋白表达,以及子代病毒的增殖。
1.11 统计学分析
各试验均平行重复3次,通过GraphPad Prism 8.0软件中-test检验方法对数据结果进行统计学分析。
2 结 果
2.1 Cas9/sgRNA载体的构建
为构建15基因敲除PK-15细胞系,针对猪源15的第二外显子,设计了两个特异性的CRISPR-Cas9 sgRNA。利用Ⅰ末端序列将sgRNA克隆到pCompass-LVKO载体上,挑取单个菌落进行DNA测序,测序结果表明,重组质粒pCompass-LVKO-sgRNA构建成功(图1),提取质粒用于后续试验。
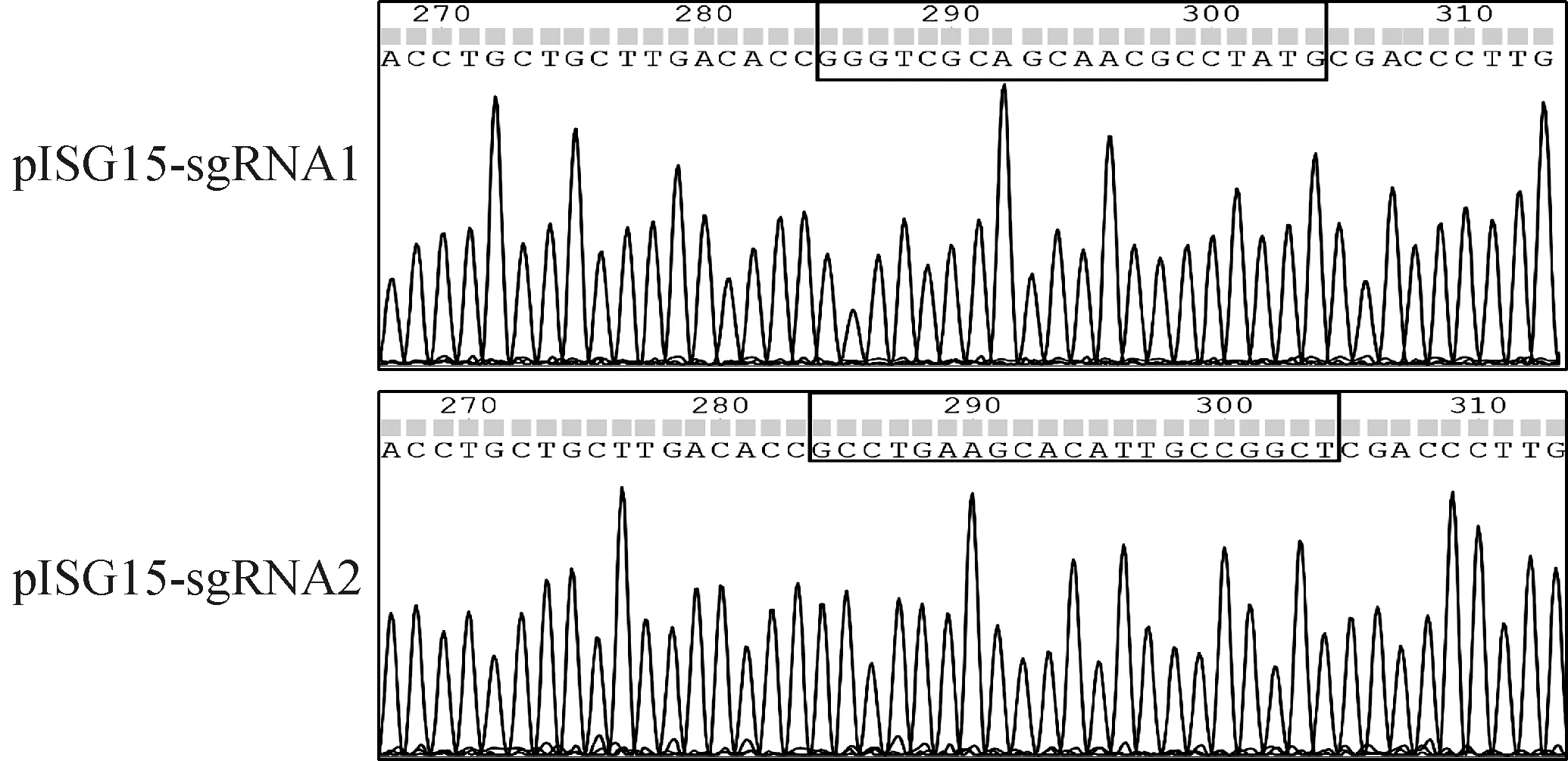
框内序列分别为sgRNA1和sgRNA2序列The sequences in box are sgRNA1 and sgRNA2 sequence respectively图1 pISG15-sgRNA测序结果Fig.1 Sequencing results of pISG15-sgRNA
2.2 PK-15敲除ISG15基因单克隆细胞系的构建
将构建成功的质粒pCompass-LVKO-sgRNA转染PK-15细胞,并以pCompass-LVKO空载体作为阴性对照。转染24 h后进行嘌呤霉素筛选,通过有限稀释法挑选单克隆。单克隆扩大培养后提取细胞总RNA,反转录为cDNA模板对15进行PCR扩增。结果显示,在619 bp处有出现一条特异性条带,初步说明该敲除细胞构建成功 (图2A)。将阳性克隆进行测序鉴定,与对照细胞的序列对比分析结果显示,sgRNA1细胞株的15基因外显子打靶区缺失20 bp碱基,sgRNA2细胞株的15基因外显子打靶区缺失8 bp碱基,选取sgRNA2 细胞株进行后续功能评价,并命名为PK15-ISG15。
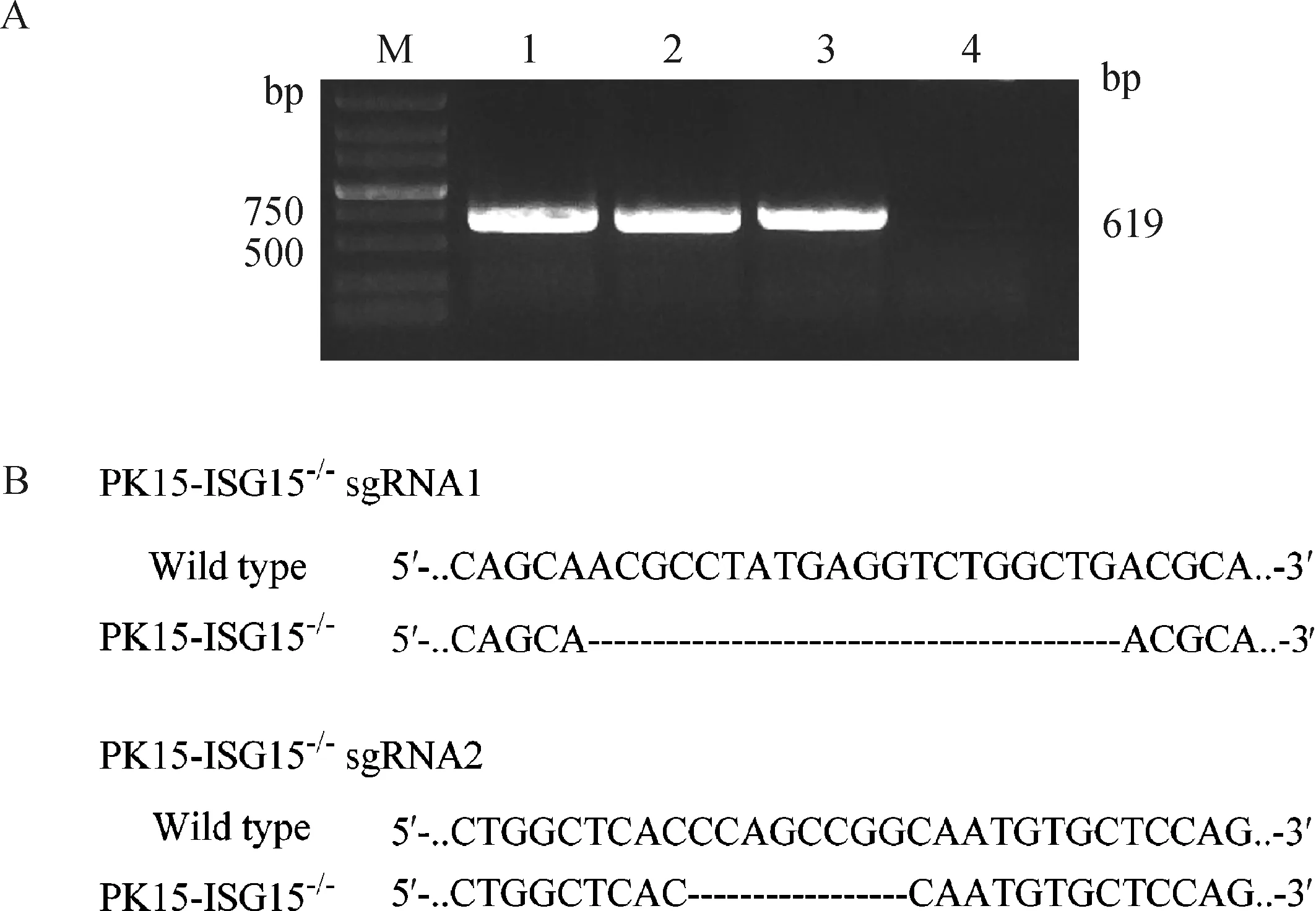
A. PCR扩增(M.DL5000 DNA相对分子质量标准;1~3. 阳性克隆细胞PCR结果;4. PK-15细胞为阴性对照);B. PK15-ISG15-/-单克隆细胞测序结果A. PCR amplification (M. DL5000 DNA marker; 1-3. The ISG15 PCR results of cell colonies; 4. PK-15 cell as a negative control); B. Sequencing results of PK15-ISG15-/- monoclonal cell图2 PK15-ISG15-/-单克隆细胞的筛选Fig.2 Screen of PK15-ISG15-/-monoclonal cell line
2.3 ISG15基因敲除效率的鉴定及对细胞增殖的影响
为验证15敲除效率,Western blot检测PK-15和PK15-ISG15细胞中ISG15蛋白的表达水平。结果表明,在PK-15细胞中可见明显的ISG15条带,而PK15-ISG15细胞中没有检测到ISG15的表达,证实PK-15细胞中15基因完全敲除(图3A)。
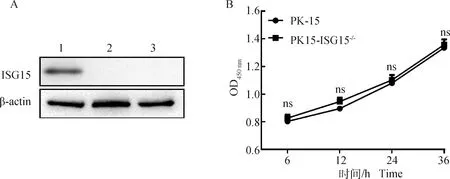
A. Western blot检测ISG15敲除效率(1.PK-15细胞;2.PK15-ISG15-/- sgRNA1;3.PK15-ISG15-/- sgRNA2);B. 敲除ISG15基因对细胞增殖无影响。ns.差异不显着A. ISG15 knockout efficiency by Western blot(1. PK-15 cell; 2. PK15-ISG15-/- sgRNA1 polyclonal cell; 3. PK15-ISG15-/- sgRNA2 polyclonal cell); B. ISG15 gene knockout can induce the replication of PK-15. ns. No significant difference图3 ISG15基因敲除细胞系的鉴定Fig.3 Identification of PK-15 cell line knockout of ISG15 gene
通过CCK-8检测敲除15基因后是否影响细胞增殖活性。结果显示,敲除15基因后PK15-ISG15细胞与PK-15细胞相比,OD波长吸光度值差异不显着(>0.05),结果表明,15基因敲除对PK15细胞的增殖没有显着影响(图3B)。
2.4 敲除ISG15基因对PRV病毒复制的影响
为了研究ISG15对PRV复制的影响,作者评估了PRV在PK-15和PK15-ISG15细胞中的复制状态。用MOI=2的PRV分别感染PK15细胞和PK15-ISG15细胞,通过间接免疫荧光检测PRV蛋白的表达,发现在感染后的不同时间点,PRV在PK15-ISG15细胞中的表达显着高于PK-15细胞中的表达(图4)。该结果初步表明,敲除15显着促进PRV复制。
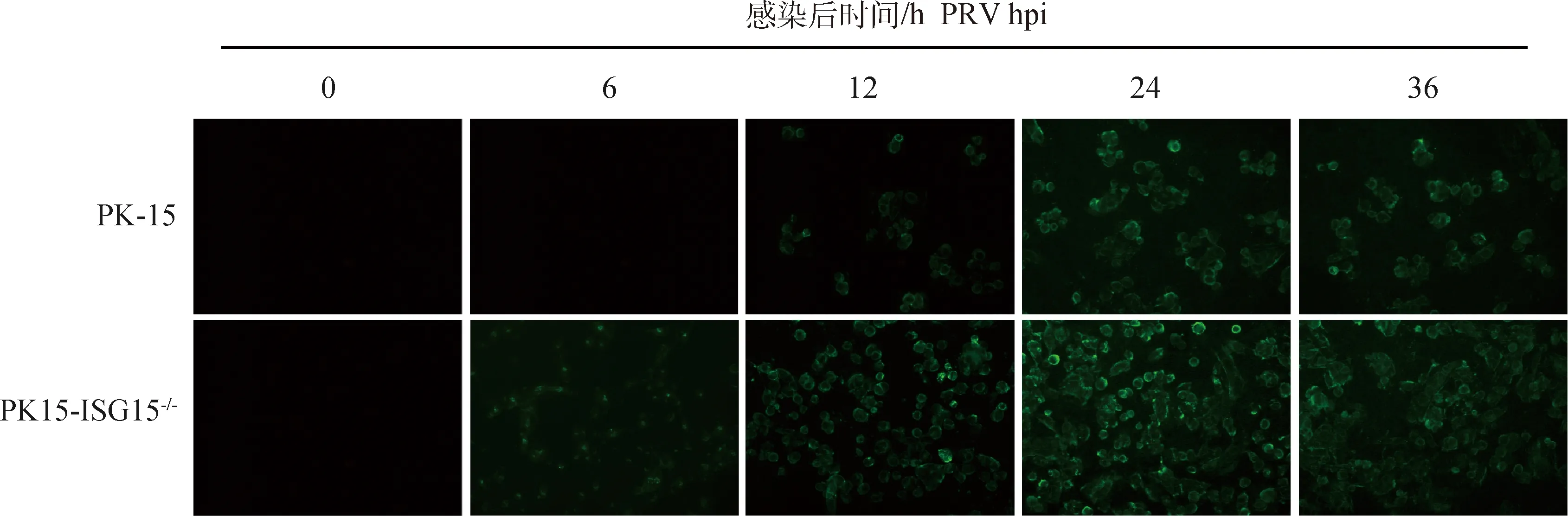
荧光显微镜观察PRV感染PK-15及PK15-ISG15-/-细胞对病毒复制的影响(80×)Effect of PRV infecting PK-15 and PK15-ISG15-/- cells on viral replication observed by fluorescence microscopy(80×)图4 敲除ISG15基因促进PRV复制Fig.4 Knockout of ISG15 gene promotes PRV replication
2.5 ISG15基因敲除对PRV不同基因及IFN-β转录水平的影响
为进一步验证15基因敲除对PRV复制的影响,利用RT-qPCR检测PK15-ISG15细胞在感染PRV后的不同时间点,PRV-0、PRV-、PRV-16的mRNA水平。与对照组PK-15细胞相比,PK15-ISG15细胞中 PRV-0、PRV-、PRV-16的mRNA水平极显着升高(<0.01;<0.001)(图5A~C),表明15基因的敲除显着促进PRV基因转录。
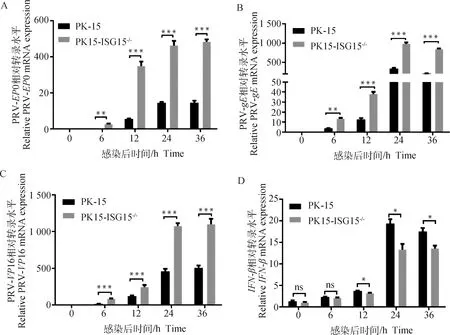
A. RT-qPCR检测感染PRV不同时间PRV-EP0基因mRNA水平;B. RT-qPCR检测感染PRV不同时间PRV-gE mRNA水平;C. RT-qPCR检测感染PRV不同时间PRV-VP16 mRNA水平;D. RT-qPCR检测感染PRV不同时间IFN-β mRNA水平。*.P<0.05; **. P<0.01; ***. P<0.001; ns. P>0.05A. PRV-EP0 gene mRNA were quantified by RT-qPCR; B. PRV-gE gene mRNA were quantified by RT-qPCR; C. PRV-VP16 gene mRNA were quantified by RT-qPCR; D. PRV-IFN-β gene mRNA were quantified by RT-qPCR. *.P<0.05; **. P<0.01; ***. P<0.001; ns. P>0.05图5 敲除ISG15基因促进PRV基因转录、抑制PRV诱导的IFN-β基因转录Fig.5 Knockdown of ISG15 promotes PRV transcription and inhibits PRV-induced IFN-β transcription
同时检测不同时间段-β的mRNA水平,结果显示,PK15-ISG15细胞中-β mRNA水平感染在0~6 h差异不显着;6~36 h时显着低于对照组(<0.05)(图5D),表明15基因敲除可以显着抑制-β的表达。
2.6 ISG15基因敲除对PRV蛋白表达的影响
相同感染复数(MOI=2)的PRV感染PK-15细胞和PK15-ISG15细胞,并在不同时间点收取细胞总蛋白,Western blot检测结果显示,PRV感染后不同时间PK15-ISG15细胞中 PRV-gE蛋白的表达量显着高于对照PK-15细胞(图6A)。PK-15细胞在感染PRV 12 h时检测到PRV-gE蛋白,而PK15-ISG15细胞在感染PRV 6 h就可以检测到PRV-gE蛋白,并且显示在PRV感染12、24、36 h的PK15-ISG15细胞检测到的PRV-gE蛋白明显高于PK-15细胞。对PRV病毒蛋白的表达进行灰度分析显示,PK15-ISG15中PRV-gE的蛋白表达极显着高于PK-15细胞(<0.001)(图6B)。以上结果表明,15基因敲除促进PRV-gE蛋白表达。

PRV gE蛋白Western blot结果(A)及灰度值分析结果(B)。***. P<0.001Western blot results of PRV gE protein(A)and gray value analysis results (B). ***. P<0.001图6 敲除ISG15基因促进PRV-gE蛋白表达Fig.6 Knockout of ISG15 gene promotes PRV-gE protein expression
2.7 ISG15基因敲除对PRV病毒滴度的影响
为研究PK-15和PK15-ISG15细胞扩增PRV子代病毒滴度的差异,用相同感染复数(MOI=2)的PRV分别感染这两种细胞,通过病毒噬斑法测定并计算PRV的病毒滴度。结果显示,PK15-ISG15细胞的空斑数多于PK-15细胞,并且每个稀释度的空斑数均高于PK-15细胞(图7A、B)。如图7C所示,随着感染时间的增加,PK15-ISG15细胞的病毒滴度显着高于PK-15细胞的病毒滴度(<0.05),并且在12 h后PK15-ISG15细胞的病毒滴度极显着高于PK-15细胞(<0.001)。结果进一步证实15基因敲除促进PRV的增殖。
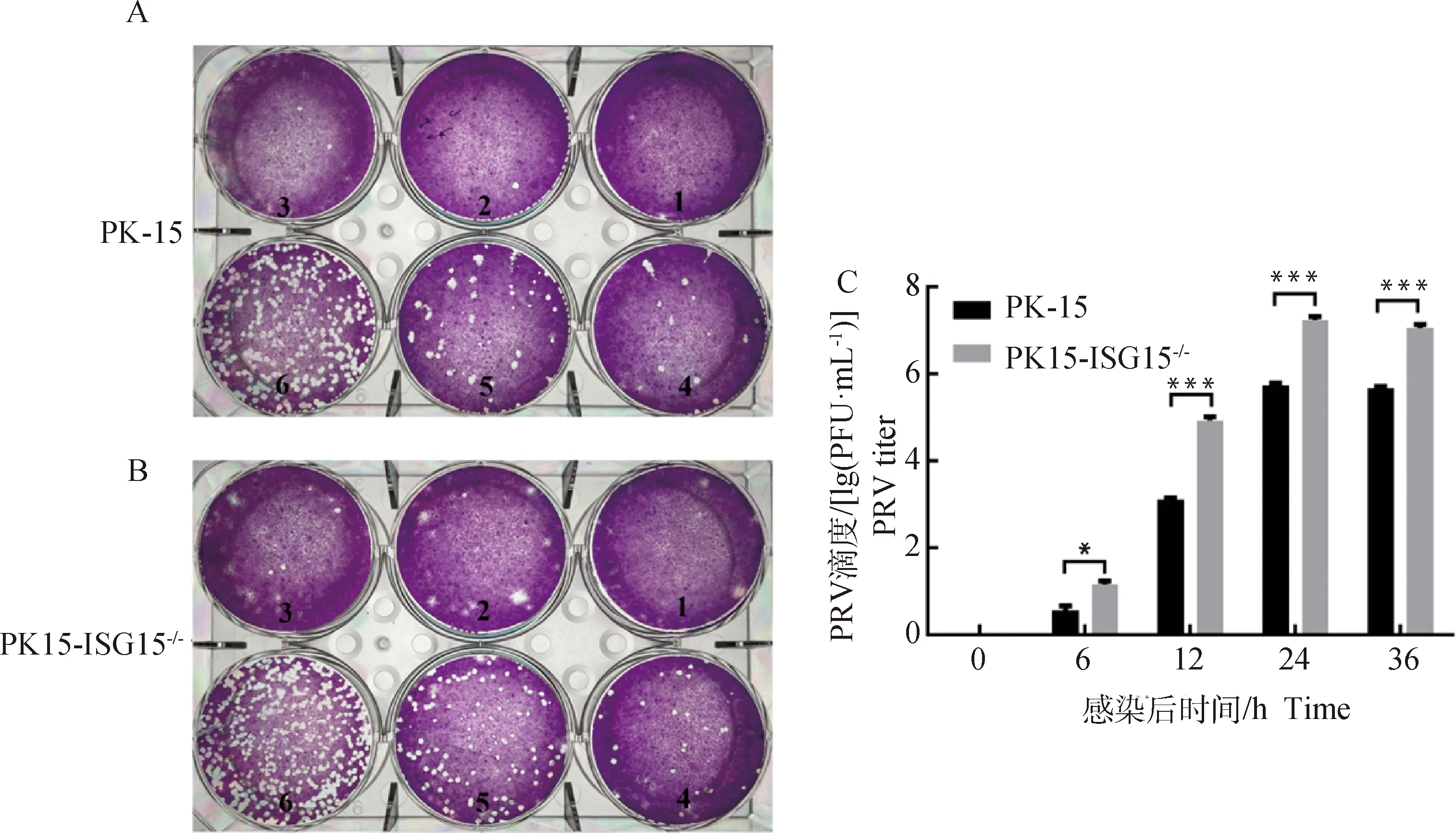
A.PRV感染24 h PK-15细胞病毒噬斑(1表示细胞对照;2~6表示病毒4倍梯度稀释,图中按照标号顺序每孔的空斑数量分别为0、1、2、14、40、153个);B.PRV感染24 h PK15-ISG15-/-细胞病毒噬斑(1表示细胞对照;2~6表示病毒4倍梯度稀释,图中按照标号顺序每孔的噬斑数量分别为0、1、7、26、94、190个);C. PK15-ISG15-/-细胞与PK-15细胞的病毒滴度。*P<0.05; ***.P<0.001A. Plaque of PK-15 cell at 24 hpi (1. Cell control; 2-6. Virus 4-fold gradient dilution, the number of plaques in each well according to the labeled order in the figure was 0, 1, 2, 14, 40 and 153, respectively); B. Plaque of PK15-ISG15-/- cell at 24 hpi (1. Cell control; 2-6. Virus 4-fold gradient dilution, the number of plaques in each well according to the labeled order in the figure was 0, 1, 7, 26, 94 and 190, respectively); C. Virus titers of PK-15 and PK15-ISG15-/- cell. *.P<0.05; ***.P<0.001图7 敲除ISG15基因促进PRV子代病毒的增殖Fig.7 Knockout of ISG15 gene promotes proliferation of PRV virus
2.8 ISG15蛋白对PRV复制的影响
为了验证ISG15是否通过抑制病毒入侵抑制PRV的复制,将纯化的ISG15蛋白与PRV于37 ℃孵育1 h后感染细胞,通过Western blot、RT-qPCR和病毒噬斑分别检测PRV的蛋白表达、转录水平以及病毒滴度,BSA与PRV孵育作为阴性对照,说明ISG15与PRV的相互作用是特异性的。Western blot检测和灰度分析结果显示,与对照组相比ISG15抑制PRV-gE蛋白的表达(<0.05)(图8A);RT-qPCR结果显示,PRV-的mRNA水平较对照组显着降低 (<0.05)(图8B);病毒噬斑结果进一步证实ISG15可以抑制PRV的复制(<0.05)(图8C)。以上结果表明,ISG15蛋白对PRV的抑制作用是通过抑制病毒侵入宿主细胞实现的。
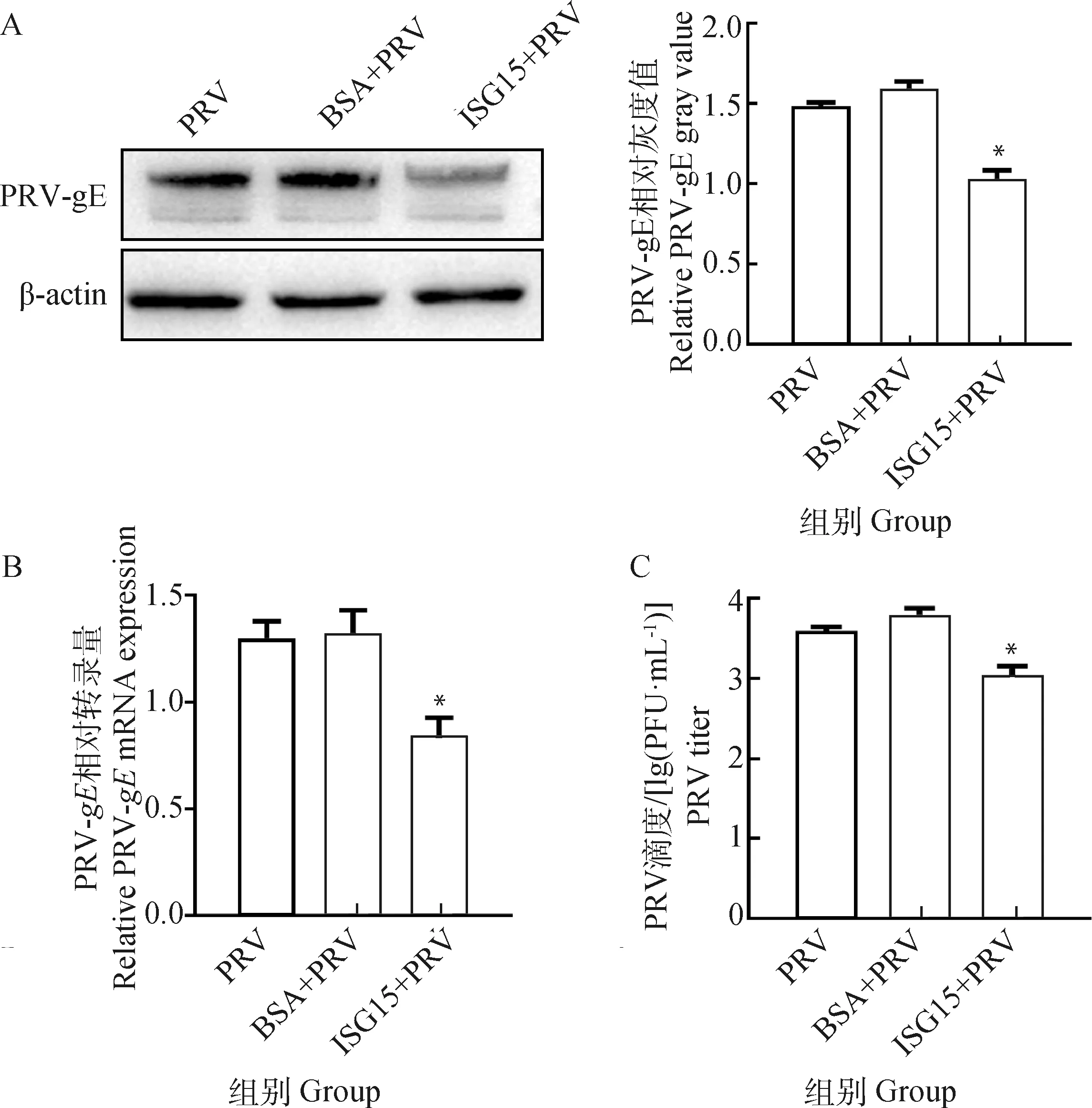
A. Western blot检测不同处理PRV感染后PRV-gE蛋白表达(左)及灰度值分析结果(右);B. RT-qPCR 检测不同处理PRV感染后PRV-gE基因的mRNA水平;C. 病毒噬斑检测不同处理PRV感染后病毒滴度。*. P<0.05A. Expression of PRV-gE protein after infection with different treatments of PRV by Western blot (left) and gray value analysis results (right); B. Expression of PRV-gE protein after infection with different treatments of PRV by RT-qPCR; C. Virus titer after infection with different treatments of PRV by plaque. *. P<0.05图8 ISG15通过抑制病毒入侵抑制PRV的复制Fig.8 ISG15 inhibits PRV replication by inhibiting viral invasion
3 讨 论
ISG15是由病原微生物IFN诱导产生的,在宿主防御和清除外源病原感染方面发挥重要作用。ISG15已被证明在病毒感染过程中发挥重要作用,既可以通过直接作用影响病毒感染的各阶段,也可以通过间接作用影响病毒感染的宿主信号通路,发挥免疫调节作用。由于作用机理的差异,ISG15在不同病原体之间甚至不同宿主物种之间的重要性可能会有所不同。对ISG15的研究有助于抗病毒药物的研发,也能为干预许多疾病的进展提供新的靶标。本试验利用近年来发展迅速的CRISPR/Cas9基因编辑技术成功构建了稳定敲除15基因的PK-15细胞系,选取sgRNA2细胞株进行功能评价,并将该细胞克隆命名为PK15-ISG15,用于研究ISG15对PRV复制的影响及作用机制。
敲低15可促进A型流感病毒(influenza virus,IAV)的复制。Nicholl等研究表明,单纯疱疹病毒Ⅰ型(herpes simplex virus-1,HSV-1)感染可以激活15、等干扰素应答基因,从而抑制HSV-1的复制。Hsiao等研究发现,过表达ISG15可以显着抑制日本脑炎病毒(Japanese encephalitis virus,JEV)的复制。本课题组前期研究发现,PRV感染PK-15细胞后引起ISG15的表达上调,过表达ISG15显着抑制PRV的复制。然而,ISG15抑制PRV复制的分子机制尚不清楚。本研究利用构建的PK15-ISG15细胞系对15基因在PRV感染过程中的作用加以探究。结果表明,15基因的敲除能够显着增加PRV基因的转录、病毒蛋白的翻译以及病毒的增殖,由此确定了ISG15对PRV具有抗病毒作用,敲除ISG15能够促进PK-15细胞中PRV的复制。
病毒入侵机体后,免疫系统可以识别病毒来源的核酸,诱导产生Ⅰ型IFN、促炎性细胞因子等。Chen等发现PRV感染过程中,ISG20在PK-15细胞中通过增强IFN-β信号抑制PRV复制。ISG15在抑制PRV复制过程中发挥关键作用,在本研究进一步发现,ISG15敲除能抑制IFN-β的表达,推测 ISG15抗PRV复制的作用可能是通过调节Ⅰ型IFN实现,这其中的具体作用机制还需进一步探究和阐明,本文构建的15敲除细胞系可以为该机制的研究提供有力工具。
4 结 论
从PK-15细胞中敲除15并通过IFA、RT-qPCR、Western blot和病毒噬斑评估了15基因敲除后对PRV复制的影响,确定了ISG15对PRV的抗病毒作用。进一步发现敲除15能够抑制IFN-β的表达,在一定程度上促进PRV的复制。本研究为进一步探讨ISG15抑制PRV复制的分子机制提供良好的生物材料。



