郭妍婷,周建民,王 晶,齐广海
(中国农业科学院饲料研究所 农业农村部动物产品质量安全饲料源性因子风险评估实验室(北京) 农业农村部饲料生物技术重点开放实验室,北京 100081)
蛋壳是禽类进化过程中为保护繁衍而形成的一种特殊结构,是蛋内物质与外界环境的交流通道,能够为胚胎发育提供物理保护,并防止微生物入侵。鸡蛋蛋壳由大约95%的碳酸钙(CaCO3)、3.5%的有机基质和1.5%的水组成[1]。钙是蛋壳矿化过程中最为关键的矿物元素,直接影响蛋壳品质[2],蛋壳中的钙主要来自饲粮和骨骼。饲料是蛋壳矿化中Ca2+的主要来源,经过消化道吸收进入体液,再由体循环运送到子宫部参与蛋壳矿化。在蛋壳矿化形成时,髓质骨破骨作用加剧,向子宫部提供了部分Ca2+。无机矿化离子[主要为Ca2+和碳酸氢根离子(HCO3-)]和有机基质蛋白相互作用并运输至子宫部矿化位点是蛋壳矿化的关键环节[3]。矿化离子在子宫部的跨膜转运主要有主动运输和被动运输。被动运输主要是靠膜两侧的离子浓度差,是无需耗能的扩散过程。Ca2+的跨膜转运系统包括跨细胞途径(主动运输)和旁细胞途径(被动运输),这一直以来是子宫部钙吸收和转运研究的重点。近年来随着对胞外囊泡研究的深入,其对子宫部Ca2+转运和蛋壳矿化的作用成为新的研究热点[4]。此外,转录组学和蛋白组学的成熟应用正鉴定出越来越多参与了蛋壳矿化的Ca2+转运相关基因和蛋白,为理解Ca2+转运在蛋壳矿化发生和调节中的作用提供了新视角[1,5-6]。因此,以蛋壳矿化期的Ca2+转运为靶点,通过营养干预的方式改善生产中蛋壳品质下降的问题具备可行性。本文综述了蛋壳矿化期子宫部Ca2+的来源和转运途径,Ca2+转运对蛋壳矿化和蛋壳品质的影响及其营养调控措施,旨在为生产中鸡蛋蛋壳品质的改善提供科学依据。
1 蛋壳的矿化
蛋壳的形成是一个周期性、时序性的动态过程,大约需要24 h。蛋壳的矿化主要发生在蛋鸡输卵管子宫部,是在子宫部分泌的有机基质的精密调控下,由多种离子参与交换和沉积,形成以CaCO3为主要组成的生物矿化物[7]。蛋壳超微结构中乳突层的形成是矿化初始阶段,一般发生在排卵后5~10 h,矿化速度相对较慢。排卵后10~22 h,卵到达子宫部中部,在子宫液充足、持续的离子供应下,蛋壳的矿化进入快速沉积期,形成超微结构中最厚的一层——栅栏层。排卵后22~24 h,进入矿化末期,形成垂直晶体层、胶护膜等,矿化停止[8-9]。
2 蛋鸡子宫部钙离子的转运
蛋壳矿化形成所需的大量Ca2+和HCO3-,并不在子宫部贮存,而是由血液持续供应[10]。因此子宫部Ca2+转运是指血液中的Ca2+跨过子宫上皮细胞,到达子宫液中蛋壳(或壳膜)矿化位点的过程。目前报道的子宫部Ca2+转运方式有三种:跨细胞途径、细胞旁途径和细胞外囊泡途径[3-4]。
2.1 钙离子的跨细胞和细胞旁转运途径
跨细胞转运是蛋鸡子宫上皮细胞Ca2+转运的经典途径。Ca2+从血液进入到子宫上皮细胞需依赖瞬时受体电位阳离子通道蛋白(TRPV6),在细胞内与钙结合蛋白d28k(CaBP-d28k)结合,完成贮存或运输。除CaBP-d28k外,子宫上皮细胞的内质网对Ca2+稳态也具有调控作用[11]。细胞内Ca2+过量时,通过肌浆网/内质网Ca2+-ATP酶(ATP13A5、ATP2A2/3)运输至内质网储存;当胞内Ca2+浓度不足时,又通过Ca2+储存库——肌醇1,4,5-三磷酸受体(ITPR1/2/3)从内质网释放补充[12]。Ca2+从子宫上皮细胞排出到子宫液的过程则需质膜Ca2+-ATP酶(plasma membrane calcium ATPase,PMCA)和Ca2+/Na+交换器的参与[13]。因此,子宫上皮细胞Ca2+的跨细胞转运主要有3步:Ca2+首先通过Ca2+通道或Ca2+转运体进入子宫上皮细胞一侧的顶膜;其次,Ca2+进入细胞质与Ca2+结合蛋白结合转运到基底侧膜;最后,Ca2+由子宫上皮细胞进入子宫液。为维持子宫上皮细胞内的离子平衡,Ca2+在子宫部转运的同时也伴随Na+、K+、Mg2+、H+和Cl-的转运[6](图1),因此涉及多种离子转运体的参与(表1),包括Na+通道(SCNN1α/2β/3γ)、Na+/K+交换器(ATP1A1、ATP1B2、NKA1N4)、K+内流通道(KCNJ2/15/16、KCNMA1、KCNH1)、Cl-通道(CLCN2/5)、H+泵(ATP6V1B2、ATP6V1C2、ATP6V1G3、ATP6V0D2)和Mg2+通道(SLC41A2/3、NIPAL1)(表1),且Ca2+转运相关基因参与蛋壳的形成,表达随蛋壳钙化阶段而发生变化[6,14]。此外,血液Ca2+也可直接通过上皮细胞之间的紧密连接蛋白进入子宫液[15],此为细胞旁途径。紧密连接蛋白包括闭合蛋白、闭锁蛋白、闭合小环蛋白等[16]。
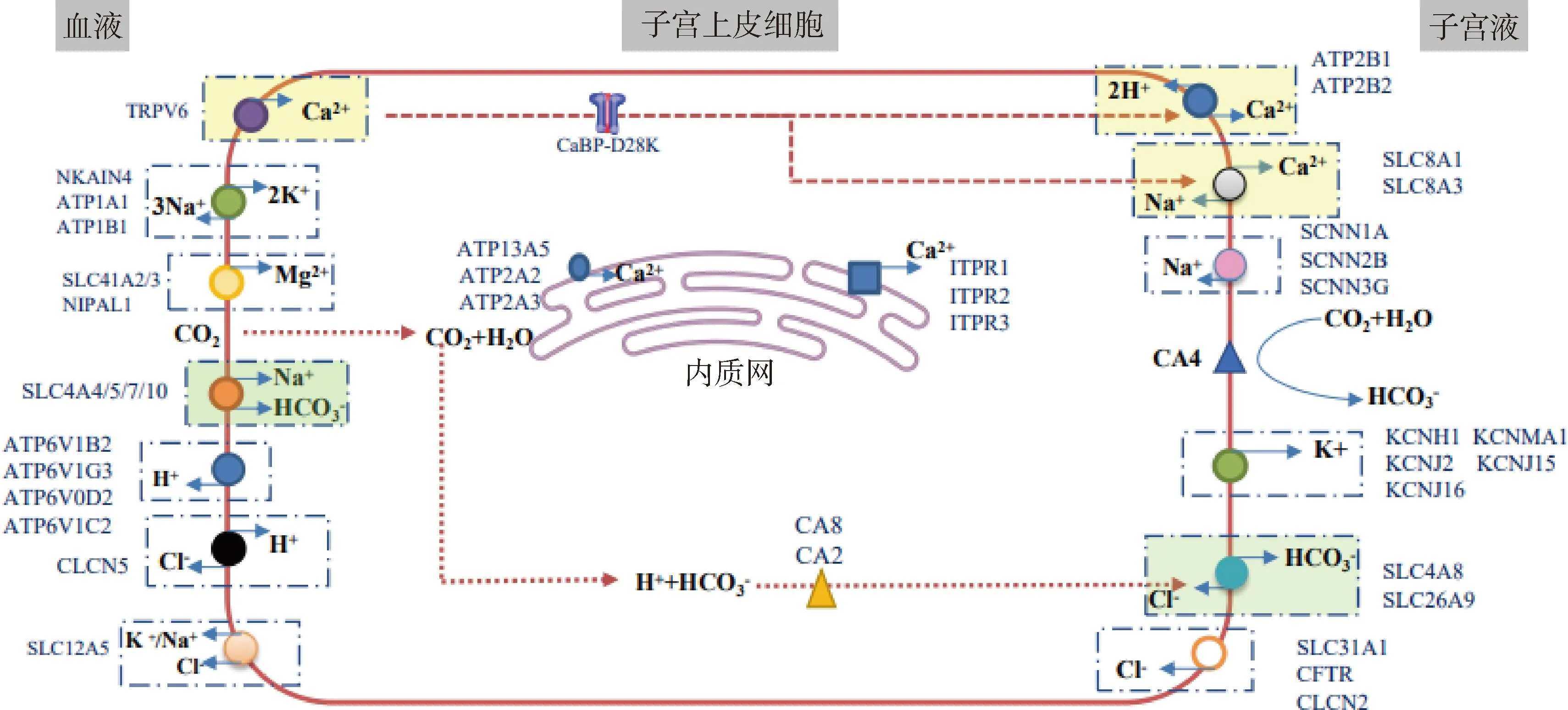
图1 蛋鸡子宫上皮细胞的离子转运(参考Broinne等[19]和Zhang等[20])Fig.1 The ion transport in uterine epithelial cells of laying hens (Referring to Broinne et al[19] and Zhang et al[20])
子宫部的HCO3-主要来自血液中的HCO3-和CO2[17]。血HCO3-依赖Na+/HCO3-共转运体——溶质载体家族4成员4、5、7和10(solute carrier family 4, member 4/5/7/10,SLC4A4/5/7/10)进入子宫上皮细胞。CO2则通过扩散进入上皮细胞,在碳酸酐酶4(CA4)的催化下与水形成HCO3-。子宫上皮细胞内HCO3-的转出则通过HCO3-/Cl-交换器——溶质载体家族4成员8(solute carrier family 4, member 8,SLC4A8)和溶质载体家族26成员9(solute carrier family 26, member 9,SLC26A9)。转出的HCO3-随后与子宫液中游离Ca2+结合形成碳酸钙前体物质,沉积到蛋壳矿化位点[18]。综上所述,蛋壳矿化是多种离子转运(Ca2+、HCO3-、Na+、H+、Mg2+、K+和Cl-)调控的协同过程,在此过程中,Ca2+和HCO3-以及有机质前体分泌到子宫液中相互作用形成蛋壳[8]。因此,这些离子转运基因和载体与蛋壳矿化和蛋壳品质有密切联系。
2.2 胞外囊泡转运途径
胞外囊泡是由脂质双分子层包绕形成的球状膜性囊泡。已发现并证实,蛋鸡子宫上皮细胞可通过分泌细胞外囊泡(extracellular vesicles,EV)转运蛋壳矿化所需的碳酸钙[4]。有研究在蛋鸡输卵管中鉴定到了胞外囊泡标志物的mRNA,并证实EV参与了蛋壳矿化[4]。Waqas等[21]利用透射电镜观察到蛋鸡的子宫上皮细胞和子宫液中存在与EV发生有关的多泡小体和包含无定形碳酸钙(ACC)的EV。Riou等[22]研究了蛋鸡子宫液中蛋白质的分子功能,发现有29种蛋白质与多泡小体的生物发生有关,并在EV中检测到热休克蛋白家族成员8、热休克蛋白家族成员90α、含缬酪肽蛋白、膜联蛋白A(annexin-A1/2/4/5,ANXA-1/2/4/5)、帕金森氏病蛋白7和苹果酸脱氢酶蛋白1等标志蛋白。根据现有证据,推断EV转运Ca2+的模式如图2所示:在蛋壳矿化开始时,子宫上皮细胞分泌EV到子宫液,EV中的ANXA形成Ca2+通道,Ca2+与ANXA上的色氨酸残基结合被转运至EV中,与CA4催化形成的HCO3-作用生成ACC,然后EV包裹着ACC运输至蛋壳矿化位点开始矿化。此过程中卵清蛋白、溶菌酶和其他EV蛋白共同作用阻止了子宫液的非特异性沉淀。有研究表明EGF样重复序列和盘状蛋白I样结构域3(EGF-like repeats and discoidin I-like domains 3,EDIL3)和重组乳脂球-EGF生长因子8(recombinant milk fat globule-EGF factor 8,MFGE8)可将含有ACC的EV靶向引导到矿化位点[23]。目前对蛋鸡子宫部EV转运途径的研究仍十分有限,EV在蛋壳矿化过程的作用和调控机制值得深入研究。
3 子宫部钙离子转运对蛋壳矿化和蛋壳品质的影响
蛋壳矿化初始期碳酸钙的平均沉积速度约为0.1 g·h-1,快速沉积期为0.33 g·h-1。子宫部Ca2+的转运涉及了多个基因及蛋白。Jonchère等[6]通过F-2α前列腺素注射使鸡蛋在蛋壳矿化前排出,对比了37个离子转运基因在矿化和非矿化期的表达情况,筛选到11个过表达的离子转运基因,包括Ca2+转运基因(CALB1、ATP2B1,2)、Na+转运基因(SCNN1G、ATP1A1)、K+转运基因(KCNJ2、KCNJ15和KCNMA1)、HCO3-转运基因(CA2、SLC26A9)、Cl-转运基因(CLCN2,5)、H+转运基因(ATP6V1B2);Zhang等[24]通过转录组筛选出13个呈时序性表达的与离子转运相关的基因,结果表明,CALB1、CA2、SCL26A9、SCNN1G、NKAIN4、CLCN2,5和KCNJ15不仅在矿化期过表达,且都在产蛋后16~20 h表达量达到最高。研究发现,TRPV6与蛋壳强度和钙含量呈正相关,而且是调控蛋壳矿化的关键通道,随着蛋壳矿化其表达量在快速沉积期达到最大[25];Khan等[26]对比了矿化初始期(排卵后5 h)和线性沉积期(排卵后15 h)的子宫转录组结果,发现与囊泡定位和分子合成相关的基因在矿化初始期上调,而与Ca2+转运及离子和分子合成、分泌活动相关的基因在线性沉积期表达上调,进一步,筛选和鉴定了影响蛋壳矿化和力学特性的离子转运基因[6,27]。高、低蛋壳强度((44.57±0.91)Nvs. (26.68±0.38)N)蛋鸡在蛋壳矿化期子宫部的转录组研究显示,内质网Ca2+转运基因(ATP13A5)和HCO3-转运基因(SLC4A1,SLC26A9)的表达水平在高蛋壳强度组中显着上调[20];Zhang等[28]比较了高、低强度蛋壳蛋鸡基因组,也鉴定出10个与Ca2+转运相关的差异表达基因,其中包括Na+/Ca2+共转运载体基因(SLC8A1)。另有研究报道,内质网Ca2+转运基因(ATP2A3、ITPR1)、Na+/HCO3-共转运基因(SLC4A5)和Na+/K+交换器基因(SLC8A3)显着影响蛋壳强度、蛋壳厚度和蛋壳重量[26];内质网Ca2+泵(ITPR2)与蛋壳强度和蛋壳厚度有关,并通过其通道开放时间和时长控制Ca2+分泌,从而调节蛋壳厚度[28-29]。可见,蛋鸡子宫部Ca2+转运相关基因的表达与蛋壳品质密切相关,并且在蛋壳矿化过程中呈时序性表达。
除上述离子转运基因,离子转运体对蛋壳矿化及品质也有重要影响。钙结合蛋白(CALB)不仅负责胞内Ca2+的贮存和运输,还具有促进CaCO3晶体沉积和形态调控的作用[30];PMCA作为子宫上皮细胞基底侧膜的Ca2+泵,在子宫部的差异表达会影响蛋壳品质[11];CA能够促进蛋壳膜上CaCO3晶体的生长并加速其沉积,在高蛋壳强度产蛋鸡体内表现出更高的活性[31]。

表1 蛋鸡子宫上皮细胞的离子转运载体及功能Table 1 Ion transpoters and their fuctions of uterine epithelial cells in laying hens
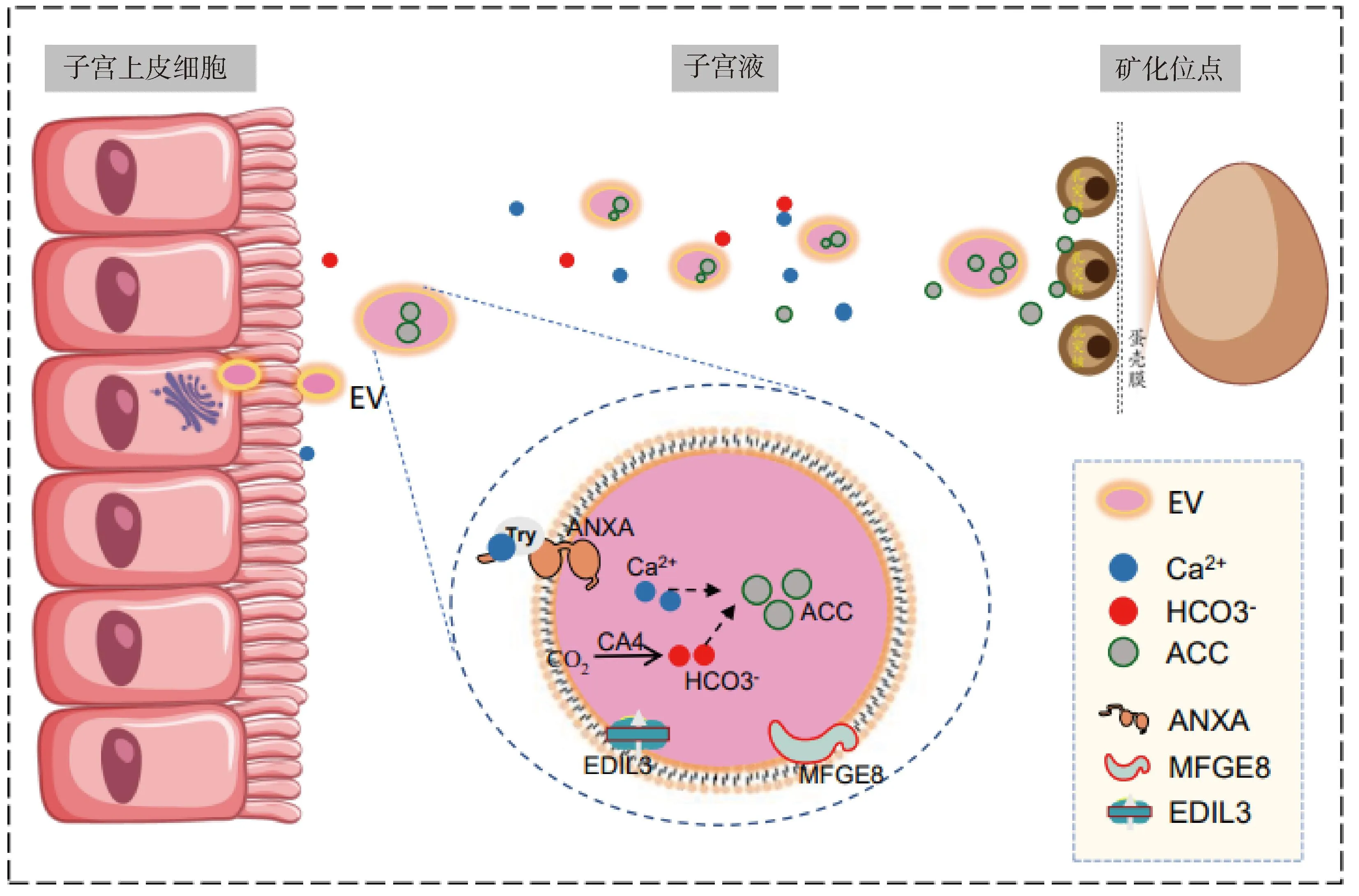
图2 蛋鸡子宫液的胞外囊泡转运(参考Stapane等[4])Fig.2 Extracellular vesicle transport in uterine fluid of laying hens (Referring to Stapane et al[4])
EV的蛋白质和脂质成分也在蛋壳矿化过程中发挥作用。ANXA是EV中种类最丰富的蛋白质,从不同类型的细胞中共分离出12种不同的ANXA,其中对ANXA5在生物矿化中的作用研究最多[34]。而EV的脂质成分磷脂酰丝氨酸(phosphatidylserine,PS)可提高ANXA5对Ca2+的亲和力,并促进CaCO3晶体的生长[33]。蛋鸡子宫部免疫荧光结果显示,EDIL3水平在蛋壳矿化起始期间显着升高[23],研究表明EDIL3和MFGE8可通过与富含Ca2+的晶体表面选择性相互作用以促进晶体成核或改变晶体形态[35]。另有研究比较了PS与ANXA5和MFGE8的结合能力,发现PS与MFGE8的亲和力更高,推测MFGE8对EV运输ACC到矿化位点起更强的调控作用[36]。综上,蛋鸡子宫部Ca2+转运是蛋壳矿化的关键过程,以子宫部Ca2+转运为调节靶点,通过营养干预的方式调节蛋壳矿化过程,可作为解决生产中多种因素(增龄、饲粮和应激水平等)诱发蛋壳品质下降问题的重要方向。
4 影响蛋鸡子宫部钙离子转运的因素
4.1 激素
Ca2+和HCO3-的离子平衡受下丘脑-垂体-性腺的激素轴调控。黄晨轩等[37]研究发现,饲粮钙水平对蛋壳品质的影响与甲状旁腺素水平有关。卵泡刺激素和黄体生成素可以促进雌激素的分泌,而蛋鸡子宫部CALB基因的表达受孕酮和雌激素的调节,孕酮通过下调子宫部中Ca2+-ATPase、CALB和PMCA的基因表达抑制蛋壳矿化[38-40],而雌激素在蛋壳矿化期间分泌增加,促进了CALB的表达[41]。Zhang等[42]研究发现,孕酮可以改变蛋壳超微结构,进而影响蛋壳品质,在蛋鸡产蛋后2 h注射孕酮,可加速乳突体的早期融合,延长矿化期,从而减少乳突层厚度并增加有效层厚度,其可能的机制是上调了矿化初始期子宫部CaBP-d28k及CA2的表达以增加子宫Ca2+转运和改善晶体结构,促进CaCO3形成与沉积;而产蛋后5 h注射孕酮却降低了蛋壳有效层厚度,可能是孕酮抑制了快速沉积期CaBP-d28k的表达使子宫部Ca2+供应不足。此外,激素水平的变化影响因素较多,如何利用营养因素通过调控激素水平间接影响Ca2+转运是研究的重点和难点。
4.2 维生素
CALB的表达与维生素D3代谢的强弱密切相关,饲粮维生素D3被摄入体内在肝和肾中转化为1,25-二羟维生素D3(1,25-(OH)2D3),1,25-(OH)2D3是类固醇激素,可促进肠上皮CALB的表达,增加钙离子的吸收和转运,进而影响蛋壳形成,提高蛋壳品质。研究发现,饲粮中添加125 μg·kg-1维生素D3和1,25-(OH)2D3增加了乳突密度和栅栏层厚度,提高了蛋壳强度,同时提高了血浆钙水平[43];350~450 IU·kg-1饲粮维生素D3水平壳显着改善蛋壳乳突层厚度以及血清钙、磷含量[44]。饲粮中维生素D3缺乏时,机体吸收钙的能力下降,蛋鸡子宫部CaCO3浓度随之降低,则会导致薄壳蛋和软壳蛋的产生[45]。然而,1,25-(OH)2D3是否调节蛋鸡子宫部CALB的表达存在争议[41,46],饲粮1,25-二羟维生素D3对子宫CALB蛋白水平没有影响[47],而体外培养蛋鸡子宫部组织中添加1,25-二羟维生素D3又可以促进CALB蛋白的表达[48]。此外,有研究表明,维生素D3可以增加上皮组织中PMCA的表达[49]。可以确定的是,蛋鸡对维生素D3或2,5-羟维生素D3的需求与钙水平一样存在上限,过量补充维生素D3虽提高了血钙浓度,但加重了蛋壳丘疹问题[50]。此外有研究发现,饲粮添加维生素A、维生素C对蛋壳品质也有一定的改善作用[44,51],但是否通过调控Ca2+转运发挥作用还需进一步研究。
4.3 矿物元素
钙、磷、铜、锌、锰、钠和氯等矿物元素是蛋壳形成所必须的,它们通过维持蛋壳矿化时子宫部的离子稳态来调控Ca2+转运,并最终影响蛋壳的矿化过程[52-55]。
4.3.1 钙、磷 钙、磷来源和水平及钙的溶解度都会对蛋壳品质产生影响,同时与Ca2+的转运有密切关系[58]。常见的钙源有石灰石、蛤壳、牡蛎壳、蛋壳等;磷的来源主要有植酸磷和非植酸磷。Lee等[56]对比了粗、细鸡蛋壳、石灰石、蛤壳和牡蛎壳对蛋壳品质的影响,结果表明粗蛋壳相较于其他组可以更好地提高蛋壳品质;粗石灰或牡蛎壳相较于细石灰石在鸡胃中停留时间长、溶解速度也较慢,可以更好地为蛋鸡供应钙,在夜间也可以维持血钙浓度[57]。饲粮中添加非植酸磷可以改善蛋壳品质[58]。随周龄增加,蛋鸡摄入矿物元素的能力逐渐下降,Ca2+转运活动减弱[59]。饲粮中钙、磷水平可直接影响产蛋鸡子宫腔内CALB1蛋白表达而改变CaCO3的沉积能力[60-61]。研究表明,70周龄产蛋鸡饲粮钙水平升高可线性提高蛋壳强度和厚度,降低蛋壳破损率[62],4%钙水平显着降低了子宫部CA活性、提高了子宫部CALB含量,从而改善蛋壳品质[63]。然而,一味地增加饲粮钙水平并非是蛋壳品质改善的“万能密码”,过高钙水平亦会引发蛋鸡子宫部Ca2+转运紊乱,造成蛋壳矿化异常[64]。Shet等[65]研究发现,饲粮中添加植酸磷降低了蛋鸡子宫部中CALB的表达,说明磷的来源和水平也会影响Ca2+转运基因,但钙的来源及溶解度对于Ca2+转运的影响还有待进一步探究。
4.3.2 锌、锰 研究发现锌源(硫酸锌和蛋氨酸锌)和水平会影响蛋鸡子宫部CA、碱性磷酸酶(ALP)、骨桥蛋白(OPN)和蛋壳基质蛋白-116(OC-116)的基因表达及血清中CA和ALP水平,调控蛋壳矿化过程[66-67]。在蛋鸭上,饲粮添加80 mg·kg-1的锌可促进鸭蛋壳矿化过程中HCO3-的生成,增加蛋壳中的钙、磷含量,并影响子宫部内CA2、SLC4A4/5和SLC26A9的基因表达[68]。锰源(硫酸锰和氨基酸锰)和水平同样影响蛋壳矿化过程,饲粮硫酸锰可线性和二次提高蛋壳钙化层中糖胺聚糖(矿化中Ca2+转运和沉积的重要调控物质)的含量,而氨基酸锰可同时提高蛋壳钙化层和基质薄膜中糖胺聚糖含量,二者通过调控蛋壳的矿化过程增加了蛋壳有效层厚度,从而改善蛋壳品质[69],转录组与PCR结合分析进一步证实了该过程还涉及锰对子宫蛋白糖基化和糖代谢的调节[70]。
4.3.3 钠、氯 电解质钠、氯也是影响蛋壳品质的重要因素。笔者团队研究发现,利用碳酸氢钠或硫酸钠替代氯化钠降低饲粮氯水平可提高钙的表观代谢率和蛋壳中钙水平,改善蛋壳有效层厚度和蛋壳强度[71]。继而以硫酸钠和碳酸氢钠分别作为钠源,发现0.71%硫酸钠(饲粮总钠0.3%)可提高蛋壳钙化层的糖胺聚糖水平,并改善蛋壳厚度和比例[72]。进一步地,笔者团队证实了硫酸钠通过上调子宫部离子转运基因SLC8A1、SCNN1α、ATP1BA和KCNMA1的表达,提高峡部ATPS、SULT、DS和CS含量,改善蛋壳矿化结构和品质(数据未发表)。因此,矿物元素在改善蛋壳品质方向上具有巨大潜力,其他微量元素如铁、硒、碘等对蛋壳子宫部Ca2+转运的影响有待进一步研究。
4.4 氨基酸
氨基酸对Ca2+转运的影响主要通过Ca2+转运相关基因表达和酶活性调控来提高蛋壳品质。研究发现,随着日粮添加精氨酸-硅酸盐-肌醇复合物水平的提高,硅酸盐和肌醇复合使用有利于提高精氨酸生物利用度[73],蛋壳钙沉积量也相应提高[74]。保持较低水平(约0.57%)的Met+Cys可以维持蛋壳厚度[75]。此外,氨基酸与矿物元素复合物似乎对改善蛋壳品质有更好的效果。饲粮中添加Zn-Met和Mn-Met可以显着增加蛋壳厚度[76];Abd El-Hack等[77]发现,日粮添加50 mg ZnO和100 mg Zn-Met可以提高蛋鸡血清中Zn水平;最新一项研究发现,在蛋鸡饲粮中添加一种蛋白质锌也可以通过提高血清Zn水品显着提高蛋壳品质[78]。进一步研究表明,饲粮添加80 mg·kg-1Zn-Met显着提高了血清钙含量和子宫CA、CaBP-d28k的基因表达,促进蛋壳矿化中Ca2+的沉积而提高蛋壳品质[67];Min等[79]研究发现饲料添加蛋氨酸羟基类似物螯合锌通过提高肝组织CA活性,促进Ca2+沉积,改善蛋壳品质。
4.5 植物提取物
近年来,植物提取物(类黄酮、精油)被发现可能通过促进钙、磷等矿物质的吸收或增强Ca2+转运能力提高蛋壳品质[80-81]。类黄酮主要通过影响机体生长激素释放激素、促黄体素、孕酮等激素水平而调控Ca2+转运过程,Xiao等[82]在饲粮中添加大豆苷元与中草药混合物,发现机体钙、磷含量、黄体生成素水平显着提高;4,7-二羟基异黄酮显着提高机体孕酮、卵泡刺激素、黄体生成素和雌激素水平,从而促进子宫中CaCO3晶体形成相关酶的产生提高蛋壳品质[83-84];桑叶黄酮可以通过提高Ca2+转运相关基因CALB、KCNA1的表达,提高蛋壳品质等[85];在患有子宫炎症的蛋鸡饲粮中添加植物精油,可以通过提高CALB1、ATP2B1、ATP2B2、SLC26A9的表达来改善蛋壳品质[86]。此外,酚类物质、牛至精油、八角茴香精油等也被证明有提高蛋壳品质的作用[87-89],但对其是否及如何参与蛋鸡子宫部蛋壳矿化期的Ca2+转运调节还有待进一步研究。综上,植物提取物对矿化调控的作用有待深入研究。
5 总结与展望
近十余年来,中外学者对鸡蛋蛋壳矿化过程的认知取得了较大进展。CaCO3作为蛋壳形成的最主要原料,子宫部Ca2+的有效运无疑是蛋壳矿化的关键环节和保证。在已知的三条Ca2+转运途径中,对跨细胞和细胞旁途径的认识已取得长足进展,而对胞外囊泡转运途径和调控机制的了解仍十分有限,具有很大的创新空间和研究价值。蛋白质组学和转录组学等技术的成熟应用使得越来越多的Ca2+转运相关基因和蛋白与蛋壳矿化过程联系了起来,这些因子对蛋壳力学特性、化学组成和微观结构的调控也正被逐渐认知,这对以Ca2+转运为切入点,从遗传或营养角度优化蛋壳矿化程序,改善蛋壳品质提供了靶点和可能。从营养的角度看,对蛋壳矿化和品质有实在或潜在调节效应的物质有很多,包括矿物质、激素、氨基酸等常规营养素和植物提取物等饲料添加剂。然而,目前对这些物质如何参与输卵管子宫部的Ca2+转运,及调节矿化过程的具体机制的研究仍不够深入,一定程度限制了其在生产上的广泛应用。笔者认为,以蛋壳矿化过程的Ca2+转运为突破口,未来值得深入开展研究的方向有二:一是探索建立成熟的蛋鸡子宫上皮细胞培养技术,助力从跨细胞和旁细胞途径探究Ca2+转运对蛋壳矿化和受外部调节的影响;二是加强胞外囊泡转运途径的深入研究,利用多种新兴的技术和仪器开展囊泡形成和分泌机制、形态特征、理化性质、功能特性和调控机制等研究,力求更深入的理解其在蛋壳矿化中的作用和调控潜力。



