谭 微,陈小兰,王楚琼,姜 泊,何继满
(南方医科大学附属南方医院消化内科,广州510515)
生长激素(GH)在机体的生长、发育、代谢和衰老等生命过程中起着重要作用;其靶器官包括肝脏、肺、肾脏、肌肉、骨骼等,其中肝脏是GH最重要的靶器官[1-2]。GH已广泛应用于临床,不再仅仅用于生长激素缺乏的治疗,新的应用已经推广到治疗人类免疫缺陷病毒(HIV)相关的消耗综合征和脂肪再分布综合征、胱氨酸蓄积症、炎症性疾病如炎症性肠病(IBD)、慢性肾脏疾病等[3-6]。然而,长期应用GH会产生各种不良反应,在不同的动物模型中,长期GH刺激会导致GH不敏感,即GH抵抗,甚至肿瘤发生等,但其机制还有待进一步的研究,此领域的进步对GH的临床使用具有重要的指导意义。
GH首先与膜蛋白受体GHR结合,启动GHR胞内区段与JAK2的结合,从而导致GHR磷酸化,进而STAT5与GHGHR-JAK2的多聚体结合后磷酸化,磷酸化的STAT5转入细胞核内,与转录因子结合,调节基因的转录。对于该信号通路调节机制的研究具有重要意义,SOCS家族蛋白是GH信号通路的重要调节因子。SOCS家族共有8个蛋白,包括SOCS 1~7及CIS。在组织培养和GH敲除的小鼠中,有过慢性GH刺激导致STAT5不敏感的报道[7]。其机制包括下调GHR含量,增加CIS或者SOCS-2的含量及促使SHP2向胞膜移位。但是,这些潜在的受体和受体后水平的研究都还未在正常小鼠中得到验证。本文以正常幼鼠为模型,研究慢性生长激素可能的负面作用及其机制。
1 材料与方法
1.1 材料 本研究使用8周龄BALB/C雄性小鼠32只,由广东省动物研究所提供,在南方医院动物房按SPF级标准饲养。重组人生长激素(北京默克药业咨询有限公司);Beckman CS-15R高速冷冻离心机(美国 Beckman公司);Bio-Rad 200/2.0电泳仪(美国Bio-Rad);硝酸纤维素滤膜(美国Millipore公司);BCA蛋白检测试剂盒(上海碧云天公司);化学发光增强剂(ECL)试 剂 盒 (美 国 millipore 公 司);P-Stat5(Tyr694)(D47E7):#4322(Cell Signaling公司);STAT5抗体 (c-17)(sc-835)、CIS 抗 体 (N19)(sc-1529)、SOCS-2(H-74)(sc-9022)、Actin(1-19)(sc-1616)均购自santa cruz公司;山羊抗小鼠-HRP IgG(101843)、山羊抗兔-HRP IgG(101964)及兔抗山羊-HRP IgG(104119)均购自北京中杉金桥公司。EMSA试剂盒(#20148)购自Thermo公司。
1.2 方法 32只小鼠共分为4组:PBS组、PBS+single rh-GH、chronic rhGH、chronic rhGH+single rhGH,每组8只。慢性生长激素刺激组chronic rhGH和chronic rhGH+single rhGH两组共16只小鼠每天按1μg/g剂量腹腔注射rhGH,对照组PBS组和PBS+single rhGH组16只小鼠每天注射等量PBS 0.1mol/L,共3周。注射完最后1次rhGH和PBS 16 h后次日8:00处死,处死前30min PBS组和chronic rhGH两组小鼠注射PBS 0.1mol/L,PBS+single rhGH 和chronic rh-GH+single rhGH按1μg/g腹腔注射单剂量rhGH。处死前使用麻醉剂苯巴比妥充分麻醉,然后快速摘取肝脏液氮速冻,再移至-80℃冰箱长期保存。蛋白信号分子的测定中,肝组织在低温情况下用含有蛋白酶抑制剂的RIPA液匀浆,蛋白离心制取后,电泳分离,采用免疫印迹法检测小鼠肝组织总蛋白中STAT5、p-STAT5 、CIS、SOCS-2的表达;凝胶迁移实验(EMSA)分析STAT5与DNA的黏附能力。
2 结 果
2.1 慢性rhGH刺激增加小鼠体质量 rhGH注射组和PBS对照组的小鼠相比,rhGH组小鼠平均增加体质量比PBS对照组小鼠多1.25g,差异有统计学意义(P<0.05)。
2.2 慢性rhGH刺激降低小鼠肝脏内基础的p-STAT5水平完成最后1次rhGH腹腔内注射16h后次日8:00,PBS组和chronic rhGH两组小鼠处死前30min注射PBS 0.1mol/L,chronic rhGH 组小鼠肝脏细胞稳定的p-STAT5水平(0.41±0.03)显着低于PBS组(0.86±0.07),差异有统计学意义(P<0.05);但肝脏内总STAT5蛋白水平无明显差异,见图1。

图1 PBS组与chronic rhGH组基础p-STAT5的表达
2.3 慢性rhGH刺激降低STAT5与DNA的结合能力 本实验采用EMSA方法检测细胞核STAT5与DNA黏附能力,chronic rhGH组STAT5与DNA结合的水平(0.78±0.03)显着低于PBS组(1.22±0.04),差异有统计学意义(P<0.05),见图2。

图2 PBS组与chronic rhGH组小鼠肝脏蛋白STAT5与DNA的结合能力
2.4 慢性rhGH刺激后单剂量rhGH刺激能显着提高PSTAT5水平 chronic rhGH组与chronic rhGH+single rh-GH组相比,chronic rhGH+single rhGH组小鼠肝脏细胞p-STAT5水平明显高于chronic rhGH组,差异有统计学意义(P<0.05),见图3。慢性rhGH刺激后,小鼠肝脏STAT5能对外源性单剂量rhGH产生应答,较处死前30min无外源单剂量rhGH刺激组chronic rhGH组,STAT5的磷酸化明显增多。
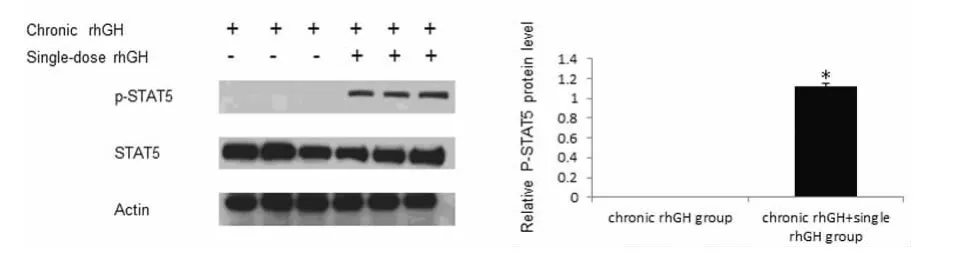
图3 慢性rhGH刺激后单剂量的rhGH刺激可以诱导大量STAT5磷酸化
2.5 慢性rhGH刺激导致STAT5对外源性rhGH不敏感PBS+single rhGH组小鼠肝脏细胞p-STAT5水平(1.19±0.04)显着高于chronic rGH+single rhGH组(0.88±0.06),差异有统计学意义(P<0.05),见图4。
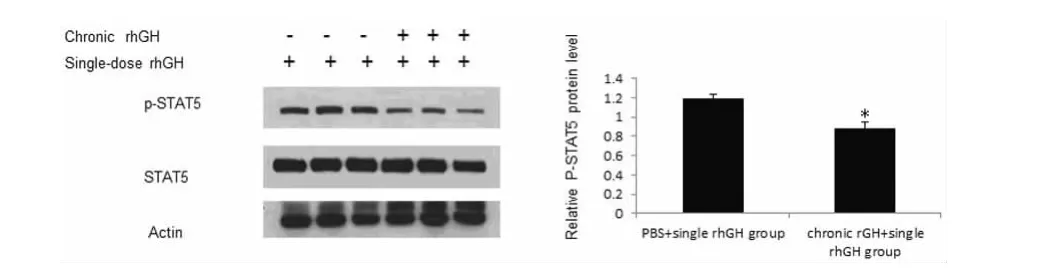
图4 慢性rhGH刺激抑制外源性rhGH引起的STAT5的磷酸化
2.6 慢性rhGH刺激升高CIS含量,SOCS-2的含量无明显改变 慢性生长激素刺激组(1.53±0.47)小鼠肝脏细胞CIS蛋白含量较对照组(0.52±0.01)明显升高,差异有统计学意义(P<0.05),但SOCS-2的含量未见明显差异(图5)。

图5 慢性rhGH刺激升高CIS含量,SOCS-2的含量无明显改变
3 讨 论
GH是机体成长、代谢等生命活动的重要调节因子,GH信号的内稳态在健康和疾病中都是很重要的。本研究显示,正常小鼠,当接受慢性rhGH刺激以后,机体同样产生对rhGH的抵抗效应。这提示了,GH抵抗的形成可能是机体对抗过量外源GH的一种普遍机制。
本研究中,GH刺激导致的小鼠肝脏内基础p-STAT5水平、由外源rhGH刺激诱导产生的p-STAT5水平以及STAT5与DNA的结合能力均降低。这与以前报道的在肝脏CWSV-1细胞系、雌性大鼠以及基因工程小鼠中的结果具有一致性[8-10]。
本课题组前期研究发现慢性rhGH刺激对内源性GH的分泌无影响[11]。这和以前报道的病理机制下的GH不敏感的情况不一样。在慢性肾脏疾病和心脏衰竭的患者中,其血液中GH浓度较健康人更高[12-13]。GH基因工程小鼠血浆中的GH浓度亦是增高[14]。
在白细胞介素-10(IL-10)结肠炎小鼠模型中发现,肝脏GHR水平下调可能与STAT5磷酸化水平的抑制有关[15]。而在本研究在正常小鼠中,当慢性生长激素刺激后,GHR水平却没有变化[11]。
当敲除SOCS-2基因后,小鼠发育异常,获得性的巨大症[16]。在尿毒症的GH抵抗模型中,部分SOCS基因的表达增加,其骨骼肌和肝脏的SOCS-2mRNA的表达大量增多,从而与GH信号通路的不敏感相联系[17-19]。然而,本研究的健康小鼠的模型中,SOCS-2的含量与对照组相比,没有明显差异。
在转基因鼠和共转染的模型中发现CIS的增加与GH信号通路不敏感相联系[20-22]。这和本研究在健康小鼠模型中的结论相一致。在健康小鼠中,慢性rhGH刺激导致的STAT5磷酸化降低是通过增加CIS的含量完成的,但CIS的作用位点和具体作用机制还有待下一步研究。
健康小鼠对慢性生长激素刺激同样产生抵抗,但其生理背景和其他病理情况不一样。相对应的,生长激素不敏感的机制也各不相同。这可能说明了,生长激素慢性刺激可以导致机体信号稳态的紊乱。也可能说明了这种生长激素不敏感是机体对抗生长激素过度刺激的一种负反馈机制。不同病理和生理情况的不敏感机制各不相同,这说明了细胞信号稳态调节的多态性。
[1] Kopchick JJ,Andry JM.Growth hormone(GH),GH re-ceptor,and signal transduction[J].Mol Genet Metab,2000,71(2):293-314.
[2] Beyea JA,Sawicki G,Olson DM,et al.Hormone(GH)receptor knockout mice reveal actions of GH in lung development[J].Proteomics,2006,6(3):341-348.
[3] Spinola-Castro AM,Siviero-Miachon AA,da Silva MT,et al.The use of growth hormone to treat endocrine-metabolic disturbances in acquired immunodeficiency syndrome(AIDS)patients[J].Arq Bras Endocrinol Metabol,2008,52(5):818-832.
[4] Grunfeld C,Thompson M,Brown SJ,et al.Recombinant human growth hormone to treat HIV associated adipose redistribution syndrome 12week induction and 24week maintenance therapy[J].J Acquir Immune Defic Syndr,2007,45(3):286-297.
[5] Gupta V,Lee M.Growth hormone in chronic renal disease[J].Indian J Endocrinol Metab,2012,16(2):195-203.
[6] Youssef DM.Resultsof recombinant growth hormone treatment in children with end-stage renal disease on regular hemodialysis[J].Saudi J Kidney Dis Transpl,2012,23(4):755-764.
[7] Gebert CA,Park SH,Waxman DJ.Regulation of signal transducer and activator of transcription(STAT)5bactivation by the temporal pattern of growth hormone stimulation[J].Mol Endocrinol,1997,11(4):400-414.
[8] Gebert CA,Park SH,Waxman DJ.Down-regulation of liver JAK2-STAT5bsignaling by the female plasma pattern of continuous GH stimulation[J].Mol Endocrinol,1999,13(2):213-227.
[9] Choi HK,Waxman DJ.Growth hormone,but not prolactin,maintains low-level activation of STAT5aand STAT5bin female rat liver[J].Endocrinol,1999,140(11):5126-5135.
[10]Choi HK,Waxman DJ.Pulsatility of growth hormone(GH)signaling in liver cells:role of the JAK2/STAT5 pathway in GH action[J].Growth Horm IGF Res,2000,10(1):1-8.
[11]王娟,高原,谭薇,等.人生长激素慢性刺激对小鼠肝细胞JAK/STAT5信号通路的影响[J].现代消化及介入诊疗,2011,16(1):28-31.
[12]Cicoira M,Kalra PR,Anker SD.Growth hormone resistance in chronic heart failure and its therapeutic implications[J].J Card Fail,2003,9(3):219-226.
[13]Rabkin R,Sun DF,Chen Y,et al.Growth hormone resistance in uremia,a role for impaired JAK/STAT signaling[J].Pediatr Nephrol,2005,20(3):313-318.
[14]Zhou Y,Xu BC,Maheshwari HG,et al.A mammalian model for Laron syndrome produced by targeted disruption of the mouse growth hormone receptor/binding protein gene(the Laron mouse)[J].Proc Natl Acad Sci USA,1997,94(24):13215-13220.
[15]DiFedele LM,He J,Bonkowski EL,et al.Tumor necrosis factor alpha blockade restores growth hormone signaling in murine colitis[J].Gastroenterology,2005,128(5):1278-1291.
[16]Greenhalgh CJ,Bertolino P,Asa SL,et al.Growth enhancement in suppressor of cytokine signaling 2(SOCS-2)-deficient mice is dependent on signal transducer and activator of transcription 5b(STAT5b)[J].Mol Endocrinol,2002,16(6):1394-1406.
[17]Schaefer F,Chen Y,Tsao T,et al.Impaired JAK-STAT signal transduction contributes to growth hormone resistance in chronic uremia[J].J Clin Invest,2001,108(3):467-475.
[18]Sun D,Zheng Z,Tummala P,et al.Chronic uremia attenuates growth hormone induced signal transduction in skeletal muscle[J].J Am Soc Nephrol,2004,15(10):2630-2636.
[19]Woelfle J,Rotwein P.In vivo regulation of growth hormone-stimulated gene transcription by STAT5b[J].Am J Physiol Endocrinol Metab,2004,286(3):393-401.
[20]Karlsson H,Gustafsson JA,Mode A.Cis desensitizes GH induced Stat5signaling in rat liver cells[J].Mol Cell Endocrinol,1999,154(1/2):37-43.
[21]Morales O,Faulds MH,Lindgren UJ,et al.1alfa,25-dihydroxyvitamin D3inhibits GH-induced expression of SOCS-3and CIS and prolongs growth hormone signaling via JAK2/STAT5system in osteoblast like cells[J].J Biol Chem,2002,277(38):34879-34884.
[22]Tollet-Egnell P,Flores-Morales A,Stavréus-Evers A,et al.Growth hormone regulation of SOCS-2,SOCS-3,and CIS messenger ribonucleic acid expression in the rat[J].Endocrinol,1999,140(8):3693-3704.



