钟晓勇,李钻芳,林如辉
(1.福建中医药大学附属人民医院,福建 福州 350004;2.福建中医药大学中西医结合研究院,福建 福州 350122)
红景天为景天科景天属(Rhodiola L.)多年生草本植物,其性味甘、涩、寒,始载于公元八世纪藏文医籍《月王药珍》,藏医药经典《四部医典》以及《神农本草经》《本草纲目》等典籍也均有记载,具有扶正固本、理气养血、润肺补肾、健脑益智和滋补强身等功效。 红景天苷(Salidroside)是从红景天中提取出的主要天然化合物,现代研究发现红景天苷等中药单体具有显着的抗缺氧、免疫调节、保护神经细胞等药理作用[1-5]。 前期研究表明红景天苷对氯化钴(cobalt chloride,CoCl2)诱导的 PC12 细胞缺氧损伤具有保护作用,其具体分子机制可能和维持线粒体结构完整性,降低活性氧(ROS)的产生以及维持雷帕霉素靶蛋白(mTOR)信号通路活性有关[6]。mTOR通路是调节细胞能量代谢的核心通路,据此推测红景天苷具有保护线粒体功能,改善能量代谢障碍的作用。 因此本研究拟观察红景天苷对缺氧所致 PC12 细胞能量代谢障碍的影响,并观察其对mTOR 及下游真核翻译起始因子4E 结合蛋白(4EBP1)激活情况,初步阐明红景天苷保护缺氧所致神经细胞损伤的分子机制。
1 材料和方法
1.1 实验材料 红景天苷(中国药品生物制品检定所);PC12 细胞株(中国科学院上海细胞生物学研究所);CoCl2(美国 Sigma 公司);DMEM 培养基、马血清和胎牛血清(美国Gibco 公司);MTT 试剂、ATP 生物素检测试剂盒、BCA 蛋白浓度测定试剂盒(上海碧云天生物技术有限公司);mTOR、P-mTOR、4EBP-1、P-4EBP-1 单克隆抗体(美国 Cell Signaling Technology 公司)。
1.2 实验仪器 SW-CJ-1FD 超净工作台(苏州安泰仪器设备有限公司);ELX 808 酶标仪(美国Bio-Tek 公司);HF212UV CO2培养箱(香港力康仪器设备有限公司);电泳仪、凝胶成像分析系统(美国Bio-Red 公司);TS-100F 型倒置显微镜(日本 Nikon 公司);垂直电泳槽、转移槽(美国 Bio-Rad 公司);Carl Zeiss LSM 710 共聚焦显微镜(德国Oberkochen公司);GloMax 20/20 发光检测仪(美国 Promega公司)。
1.3 细胞培养和分组
1.3.1 细胞培养 PC12 细胞置于10%马血清、5%胎牛血清、1%双抗(青霉素/链霉素)的完全DMEM培养基中正常培养,置于37 ℃、5% CO2的培养箱中孵育。 待细胞生长至80%~90%汇合时按1∶3 进行传代。
1.3.2 细胞接种 96 孔板:取生长良好、对数生长期的PC12 细胞,用0.25%胰酶消化后接种于96 孔培养板,每孔细胞为 1×104个,培养液 100 μL。 6 孔板:取生长良好、对数生长期的PC12 细胞,用0.25%胰酶消化后接种于96 孔培养板,每孔细胞为1×106个,培养液2 mL。
1.3.3 分组和药物干预 根据干预条件分为对照组、模型组、红景天苷组。 根据前期研究[6]结果,400 μmol/L CoCl2干预 12 h 可以使 PC12 的细胞活力下降到50%左右[(54.8±4.9)%],因此模型组继续使用超纯水稀释 CoCl2(0.22 μm 无菌过滤)至 400 μmol/L,并干预12 h 诱导缺氧损伤细胞,而对照组不用CoCl2干预。 红景天苷组在模型组的条件下,使用超纯水稀释(0.22 μm 无菌过滤)红景天苷浓度至30、60、90 μmol/L,每一浓度检测 6 个复孔,MTT 法确定红景天苷最佳干预浓度并进行后续实验。
1.4 MTT 法检测细胞活力 各组干预结束后,每孔加入 5 g/L MTT 10 μL 继续培养 4 h,小心吸弃孔内液体,加入DMSO 150 μL,置微孔振荡,待紫色结晶完全溶解后,采用ELX 800 酶标仪于570 nm 处检测,计算细胞活力。
1.5 ATP 含量检测 吸除培养液,按照6 孔板每孔加入200 μL 裂解液的比例(即相当于细胞培养液量2 mL 的1/10)加入裂解液裂解细胞;裂解后4℃12 000×g 离心5 min,取上清,按试剂盒说明书进行后续的测定,应用化学发光仪(luminometer)测得各组相对光单位(relative light unit,RLU),ATP 含量通过计算各组RLU 值和对照组的百分比得出。
1.6 Western blot 检测 mTOR 及 4EBP1 磷酸化情况 收集细胞,提取蛋白并用BCA 定量,每组取50 μg 蛋白,用15%凝胶SDS-PAGE 电泳,考马斯亮蓝染色标记蛋白质分子;然后转移至PVDF 膜上,于封闭液中进行封闭;经封闭洗涤后加入相应的单抗(1∶1 000 稀释),4 ℃过夜孵育,洗涤,增强化学发光法(ECL)显色,凝胶成像分析系统成像。 实验结果用Image Lab 进行半定量分析。
1.7 统计学方法 数据采用SPSS 22.0 统计软件处理。 计量资料符合正态分布的以表示,2 组比较采用t 检验,多组间比较采用单因素方差分析。
2 结 果
2.1 红景天苷对CoCl2诱导缺氧条件下PC12 细胞活力下降的影响 MTT 法检测结果见图1,提示不同浓度的红景天苷均能够显着缓解CoCl2所致细胞活力的降低(P<0.05)。 其中在 90 μM 浓度干预下能够达到最大细胞活力(61.6±4.4)%,提示红景天苷能够显着改善CoCl2所致的细胞缺氧损伤,对PC12 有保护性作用。 在后续实验中红景天苷组均使用90 μM 浓度红景天苷干预处理。

图1 3 组对CoCl2 所致PC12 细胞活力下降的影响
2.2 红景天苷对CoCl2诱导缺氧下PC12 细胞缺氧损伤的形态学结构影响 光镜结果显示:对照组PC12 细胞铺满瓶底,细胞形态规则完整,呈多边形整齐排列,细胞质均匀透亮;模型组贴壁细胞数量明显减少,细胞皱缩、变小、变圆,部分细胞形态不完整,胞质中可见少量颗粒状物质;而红景天苷组可部分改善CoCl2所致的PC12 细胞损伤形态学结构变化。 以上结果进一步验证红景天苷对CoCl2所致PC12 细胞缺氧损伤具有保护作用。 见图2。

图2 3 组对CoCl2 所致PC12 细胞缺氧损伤的形态学结构影响(×200)
2.3 红景天苷对CoCl2诱导缺氧下PC12 细胞ATP含量变化的影响 ATP 含量检测结果显示:与对照组比较,CoCl2会导致PC12 细胞ATP 含量显着下降(43.9±22.7)%(P<0.05),而红景天苷干预可以显着提高Col2诱导缺氧条件下PC12 细胞的ATP 含量(88.1±8.93)%(P<0.05)。 见图 3。
2.4 红景天苷对 CoCl2诱导缺氧下 PC12 细胞mTOR-4EBP1 通路活化的影响 红景天苷可以促进mTOR 以及下游蛋白4EBP1 的磷酸化水平,见图4 A;且3 组灰度值比较有统计学意义(P<0.05),见图4 B、图4C。 提示红景天苷可以在一定程度上激活mTOR-4EBP1 通路。

图3 3 组对CoCl2 诱导缺氧下PC12细胞ATP 含量变化的影响
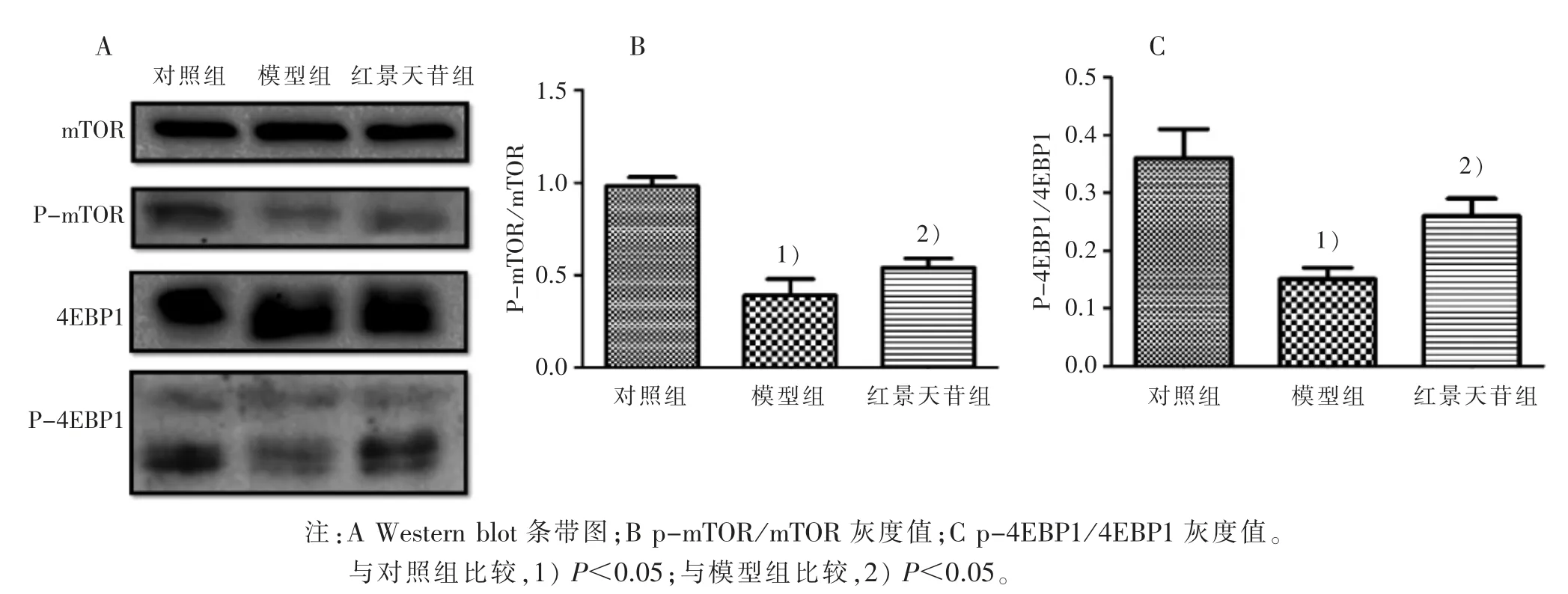
图4 3 组对CoCl2 诱导缺氧下PC12 细胞mTOR-4EBP1 通路活化的影响
3 讨 论
脑局部血液循环障碍所导致的缺血/缺氧是脑卒中发病的重要病理因素,而且由于中枢神经细胞对缺氧最为敏感,神经细胞缺氧条件下触发的氧化应激反应可以导致细胞器损伤。 其中线粒体作为细胞能量代谢的中心,是对缺氧损伤极为敏感的一类细胞器,缺氧条件下往往会导致线粒体受损。前期研究表明CoCl2干预后PC12 细胞出现明显的缺氧损伤甚至细胞凋亡的超微结构改变(核仁消失,染色质浓缩、边聚,以及线粒体肿胀、裂解、线粒体嵴消失等),而红景天苷能够减轻CoCl2所致的细胞缺氧损伤及超微结构改变,其具体机制可能和维持线粒体结构的完整性、维持mTOR 相关通路活性有关[6]。但目前红景天苷基于mTOR 相关通路改善缺氧所致细胞能量代谢障碍的作用未见报道,故本研究拟观察红景天苷对缺氧所致的PC12 细胞能量代谢障碍的影响,探讨红景天苷保护缺氧所致神经细胞损伤的分子机制。
线粒体是细胞内氧化磷酸化和合成ATP 的主要场所,mTOR 作为细胞营养感应和能量调节因子,是调控细胞的新陈代谢以及细胞周期进程和细胞生长的核心通路。目前在脑血管疾病中以mTOR 信号通路为靶点的治疗策略受到越来越多关注[7-8]。 研究表明抑制mTOR 能降低细胞内ATP 水平,造成线粒体功能损伤,糖酵解功能紊乱。 目前认为 mTOR被抑制后优先抑制细胞内编码必需核编码蛋白,包括复合物V 和线粒体转录因子mRNA 的翻译来调控上述代谢过程[9]。 mTOR 能刺激 mRNA 的翻译和其他合成代谢过程,从而调控线粒体的活性以及生物合成[10];mTOR 能够磷酸化其下游靶蛋白包括4EBP1 并启动翻译。 Morita 等[10]研究表明:4EBP1是mTOR 影响线粒体生物合成和功能主要的调节因子,通过调节mTOR-4EBP1 通路活化可以调控线粒体转录因子,调节线粒体合成以及细胞内ATP 水平。
基于上述研究结果,本实验利用CoCl2诱导建立PC12 细胞缺氧损伤模型,在诱导产生缺氧损伤模型的同时,给予不同剂量红景天苷干预,结果表明:红景天苷能够上调缺氧所致的PC12 细胞活力下降(P<0.05),其中在 90 μmol/L 浓度干预下能够达到最大细胞活力(61.6±4.4)%,提示红景天苷能够显着改善CoCl2所致的细胞缺氧损伤,对PC12 有保护性作用;同时红景天苷干预可以显着提高CoCl2诱导缺氧条件下PC12 细胞的ATP 含量,提示红景天苷可以改善CoCl2所致的细胞能量代谢障碍;根据Western blot 检测结果,进一步提示景天苷可以在一定程度上激活mTOR-4EBP1 通路。 综上,红景天苷可能通过激活 mTOR-4EBP1 通路,从而提高ATP 含量,改善缺氧所致的能力代谢障碍,减轻缺氧所致的PC12 细胞损伤,显示红景天苷在治疗缺氧所致神经细胞损伤中具有一定的临床应用价值。但mTOR 作为体内调控多种生物功能的核心通路,具有复杂的多通路调控网络,其具体作用机制仍需进一步阐明。



