饶志坚 郑莉芳 于涛 史仍飞
1 上海师范大学体育学院(上海 200234)
2 国家体育总局体育科学研究所(北京 100061)
3 上海大学体育学院(上海 200444)
4 上海体育学院运动科学学院(上海 200438)
运动是降低心血管疾病的发病率和死亡率的重要途径,此外,运动也是心脏康复中的重要内容[1-3]。除了这些效果外,运动还可直接促进心脏发生生理性重塑。这种为了匹配增加的工作负荷,由长期运动导致的心脏体积变大的现象通常称为“运动员心脏”,其特征为心肌细胞变大、新生血管增多、心功能增强或不变[4]。运动员心脏是为了适应运动而产生的一种生理性变化,它与一些疾病(如高血压、心肌梗死等)导致的病理性心肌肥大是截然不同的,病理性的心肌肥大通常伴随着心功能下降[5,6]。尽管在结构上生理性心肌肥大和病理性心肌肥大有相似的心肌细胞体积变大,然而在细胞分子层面上两者在上游信号通路的激活和下游转录反应是不同的[5,6]。为了揭示两者不同的发生机制,诸多学者通过测序、微阵列等手段探究了生理性心肌肥大和病理性心肌肥大发生过程中心肌基因表达的差异[7,8],以寻找潜在的干预靶点来促进心脏健康。运动虽然可以诱导生理性心肌重塑,但是不同的运动方式对心肌重塑的影响有所不同。类似地,我们也可以通过对比分析不同运动方式对心脏重塑过程中基因表达的差异,来揭示不同运动方式对心肌重塑具有不同效果的潜在机制。
在实验研究过程中,主要采用三种运动方式来诱导生理性心肌肥大:游泳运动、跑台运动和自由转轮运动。跑台运动的优势在于可以调节运动强度、坡度和运动时间来形成多种运动方案,研究报道依据运动方案的不同,跑台运动可使大鼠和小鼠的心肌质量增加5%~24%[9-11]。此外,至少需要经过3~4周的跑台运动才可使心肌质量显着增加。游泳运动是将小鼠或大鼠放在泳池中,通过调节游泳时间和负重来调节运动强度。现有文献报道,游泳运动可使心脏质量增加30%~50%[12-14],总体上效果要优于跑台运动。跑台和游泳运动是一种强迫性运动,因此,这两种运动可能并不是单纯的生理因素导致心肌生长,还有可能存在部分的应激因素导致心肌生长。故而有些学者会采用自由转轮运动来诱导生理性心肌肥大,文献报道经过3~4周以上的转轮运动后,小鼠和大鼠的心肌质量可增加5%~20%(依据饮食、性别、年龄等因素的不同)[15,16]。值得注意的是,研究报道自由转轮运动并不会因增加运动时间和负荷而进一步增加心肌质量[17]。可见,三种运动方式在诱导生理性心肌肥大的效果上是有所不同的。
鉴于上述,为了进一步揭示不同运动方式导致不同的运动心脏重塑的因素,本研究对不同运动方式诱导的心脏重塑过程中基因表达的情况进行分析,以期更加深入地了解运动诱导生理性心肌重塑的发生机制,为实践运动促进心脏健康提供理论依据。
1 研究方法
1.1 数据下载与处理
在基因表达数据库(gene expression omnibus,GEO)中检索运动心脏重塑相关数据集,筛选条件为运动干预时间一样、研究对象的种属及性别相同,整理发现仅有雄性小鼠游泳运动(swimming,SW)和雄性小鼠自由转轮(wheel running,WR)两种运动方式干预的数据集符合条件。因此,下载两组数据GSE77(游泳运动)和GSE36330(自由转轮)后,从两组数据中分别提取小鼠游泳3 周组及其对照组,以及小鼠自由转轮运动3 周组及其对照组,然后对数据进行标准化。如果有多个探针映射到同一个基因中,则用平均表达值来表示该基因的表达值。
1.2 筛选差异表达基因
在Rstudio中使用limma包分别处理两组数据得到游泳运动和自由转轮运动后差异表达的基因,采用t检验筛选差异表达的基因,筛选标准为P<0.05,且log2 fold change> Mean(log2 fold change)+ 2SD(log2 fold change)。然后根据log2 fold change 的值进行排序,分别选取最大的100个上调基因和100个下调基因绘制热图。同时,将所有差异表达的基因用火山图进行可视化。
1.3 GO和KEGG功能富集分析
采用R studio 中的enrichplot 包和ggplot2 包分别对两种运动后小鼠心肌中差异表达的基因进行GO(gene ontology)和KEGG(kyotoencyclopedia of genes and genomes)富集分析。P<0.05 代表差异有统计学意义。对游泳后小鼠心肌差异基因的GO 富集结果用条形图可视化,并展示差异基因与富集结果的关系图;对自由转轮运动后小鼠心肌差异基因的GO 富集结果用气泡图可视化,并展示差异基因与富集结果的关系图。
1.4 对比两种运动后心肌差异基因
对比游泳运动后小鼠心肌中上调及下调基因与自由转轮运动后小鼠心肌中上调及下调基因是否有交集,在R studio 中采用VennDiagram 包绘制韦恩图,并展示所有有交集的基因名称。
1.5 基因互作分析
采用R studio 中的STRINGdb 包对不同运动方式后小鼠心肌中的差异基因进行互作分析,并用环形图展示网络互作关系,互作得分阈值设为400,连接点数大于5 则在图中显示基因名。
1.6 基于差异基因分析转录因子
本研究在Rstudio 中采用RcisTarget 包分别对游泳和自由转轮运动后小鼠心肌中差异基因的转录因子进行富集分析。
2 结果
2.1 游泳运动后小鼠心肌中差异表达的基因
游泳运动的数据通过limma 包分析后,根据log2 fold change值展示基因表达热图(图1a)。随后按实验方法所述筛选差异基因,经计算得出cutoff log2 fold change=1.3,因此,游泳组差异表达基因的筛选标准是P<0.05 且log2 fold change>1.3。根据这一筛选标准,本研究共发现游泳运动后心肌中差异表达的基因有275个,其中129个基因上调,146个基因下调(图1b)。
2.2 自由转轮运动后小鼠心肌中差异表达的基因
自由转轮运动的数据通过limma 包分析后,根据log2 fold change值展示基因表达热图(图1c)。随后按实验方法所述筛选差异基因,经计算得出cutoff log2 fold change=0.36,因此,自由转轮组差异表达基因的筛选标准是P<0.05 且log2 fold change>0.36。根据这一筛选标准,本研究共发现自由转轮运动后小鼠心肌中差异表达的基因有856个,其中559个基因上调,297个基因下调(图1d)。
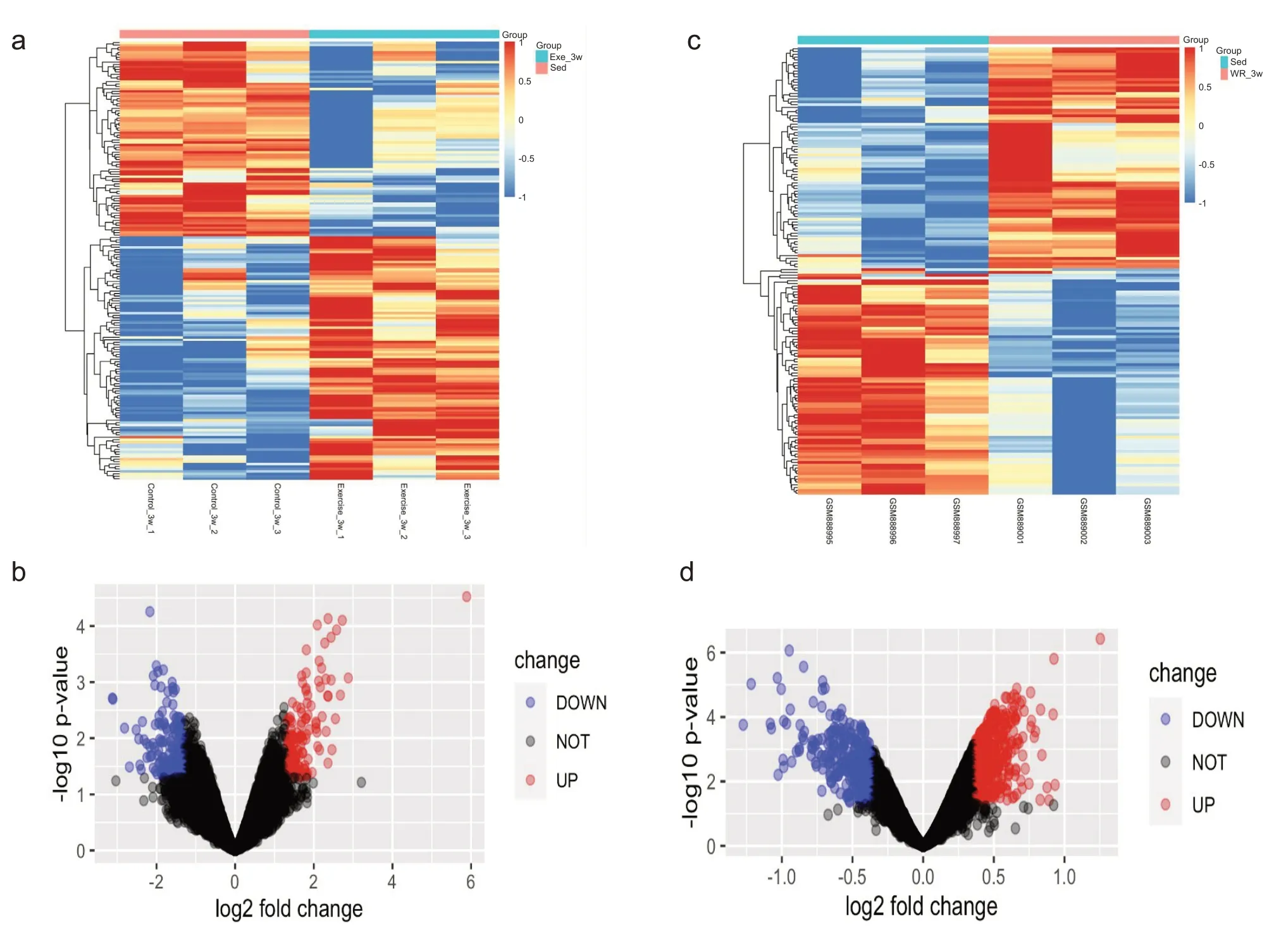
图1 运动后心肌中的差异基因
2.3 游泳运动后小鼠心肌差异基因的GO和KEGG 功能富集分析
随后本研究对游泳运动后小鼠心肌中275 个差异基因进行了GO 功能富集分析,其中细胞组分(cellular component,CC)富集结果主要是受体复合物、磷脂酰肌醇3 激酶(phosphoinositide 3-kinase,PI3K)复合物、突触部分、海马CA3区苔藓纤维突触和远端突触等(如图2a所示),差异基因与CC富集结果的关系图如图2b所示。生物过程(biological process,BP)富集结果主要涉及:血管生成的调节及其负向调节、血管形态发生的负向调节和血管发育的调节及其负向调节(如图2c所示),差异基因与BP 富集结果的关系如图2d 所示。分子功能(molecular function,MF)富集结果主要涉及:受体配体活性、离子受体活性、蛋白磷酸化氨基酸结合、磷酸蛋白结合和糖磷酸酶活性等(如图2e所示),差异基因与MF富集结果的关系图如图2f所示。
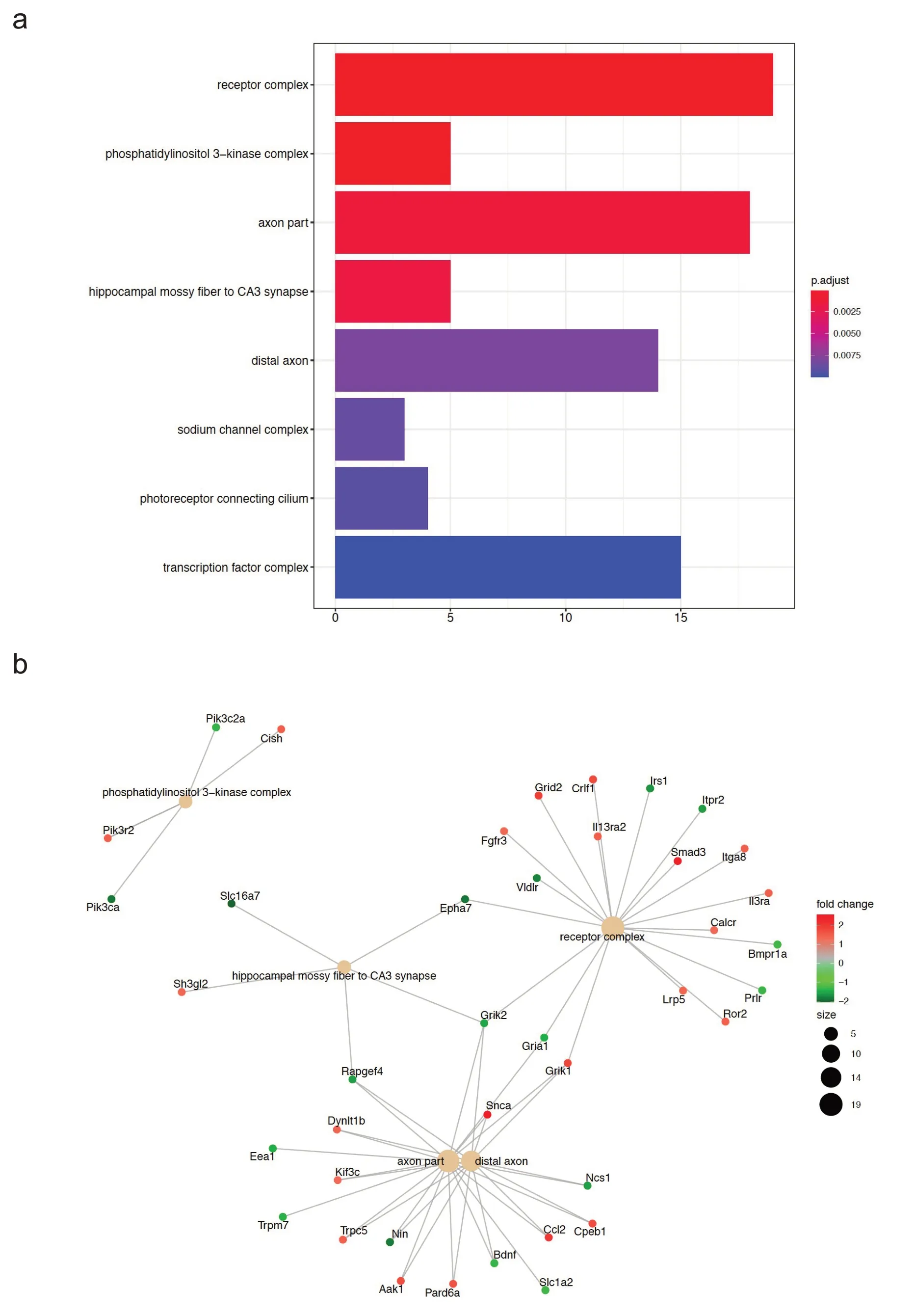
图2 游泳运动后心肌中差异基因的GO功能富集分析
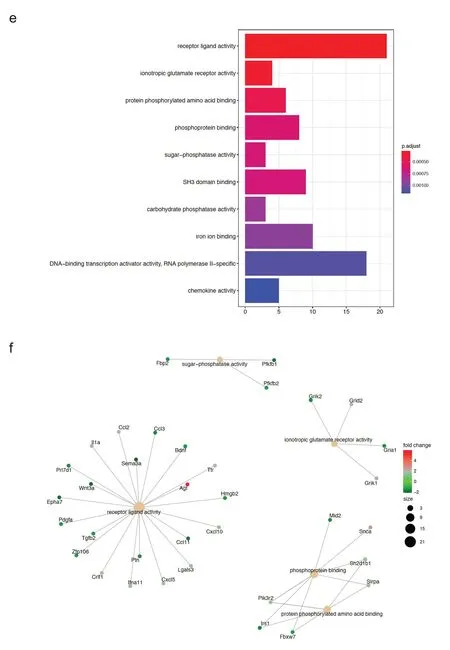
图2 游泳运动后心肌中差异基因的GO功能富集分析(续)
此外,KEGG富集结果分析主要富集在鞘糖脂合成-神经节系、AMP 活化蛋白激酶(AMP-activated protein kinase,AMPK)信号通路、风湿性关节炎、肿瘤坏死因子(tumor necrosis factor,TNF)信号通路和癌症相关microRNAs 等(如图3a 所示),差异基因与KEGG富集结果的关系图如图3b所示。
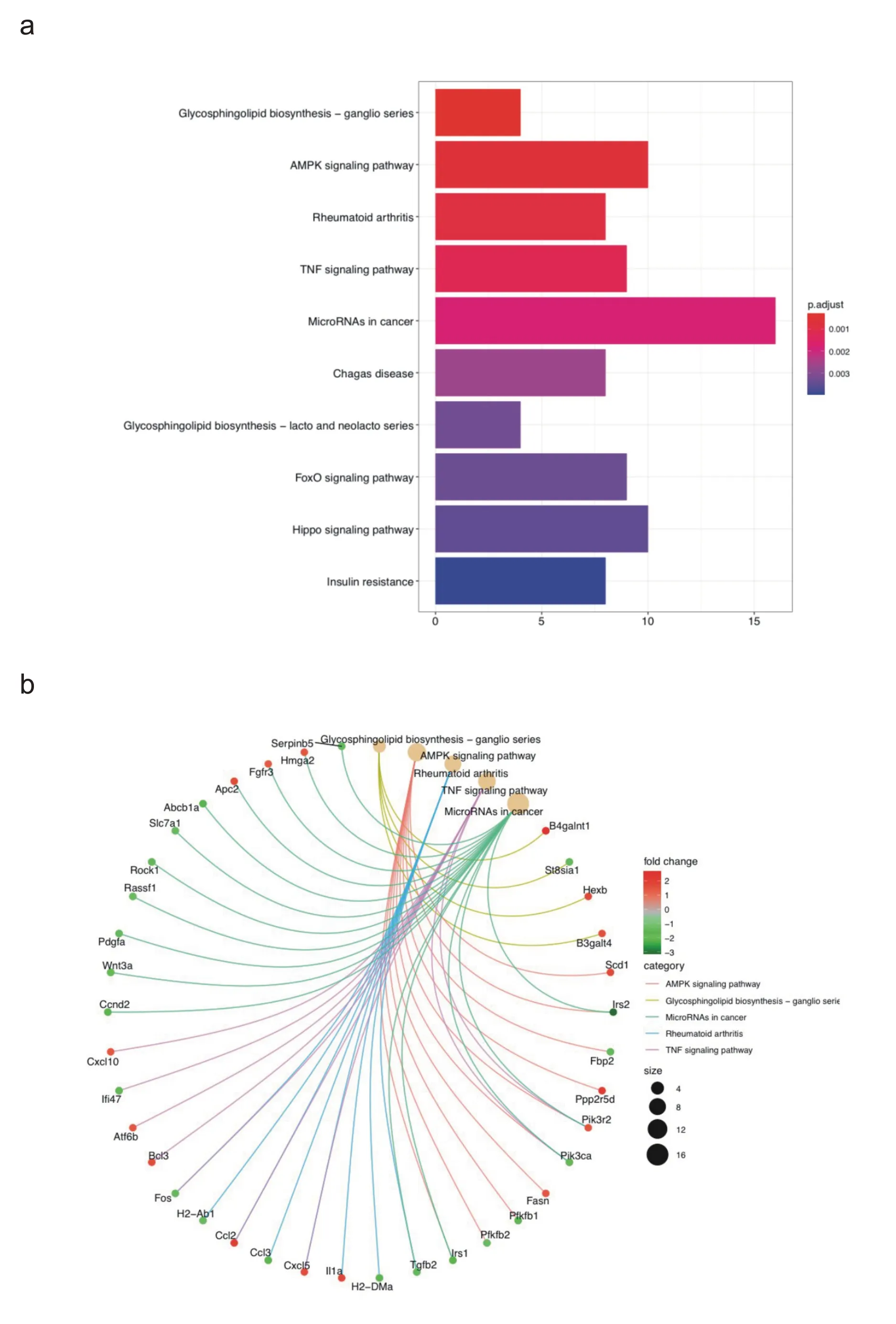
图3 游泳运动后心肌中差异基因的KEGG通路富集分析
2.4 自由转轮运动后小鼠心肌差异基因的GO 和KEGG功能富集分析
本研究对自由转轮运动后小鼠心肌中856 个差异基因进行了GO 功能富集分析,其中CC 富集结果主要是:细胞前缘、核斑点、丝氨酸/苏氨酸蛋白激酶复合物、细胞周期蛋白依赖性蛋白激酶全酶复合物和MCM复合物等(如图4a所示),差异基因与CC富集结果的关系如图4b 所示。BP 富集结果主要涉及:病毒反应、病毒防御反应、干扰素g反应、干扰素b反应和干扰素b细胞反应等(如图4c所示),差异基因与BP富集结果的关系如图4d 所示。MF 富集结果主要涉及:GTP 酶活性、多肽调节因子活性、增强子结合、金属蛋白酶抑制因子活性和死亡结构域结合等(如图4e所示),差异基因与MF富集结果的关系如图4f所示。
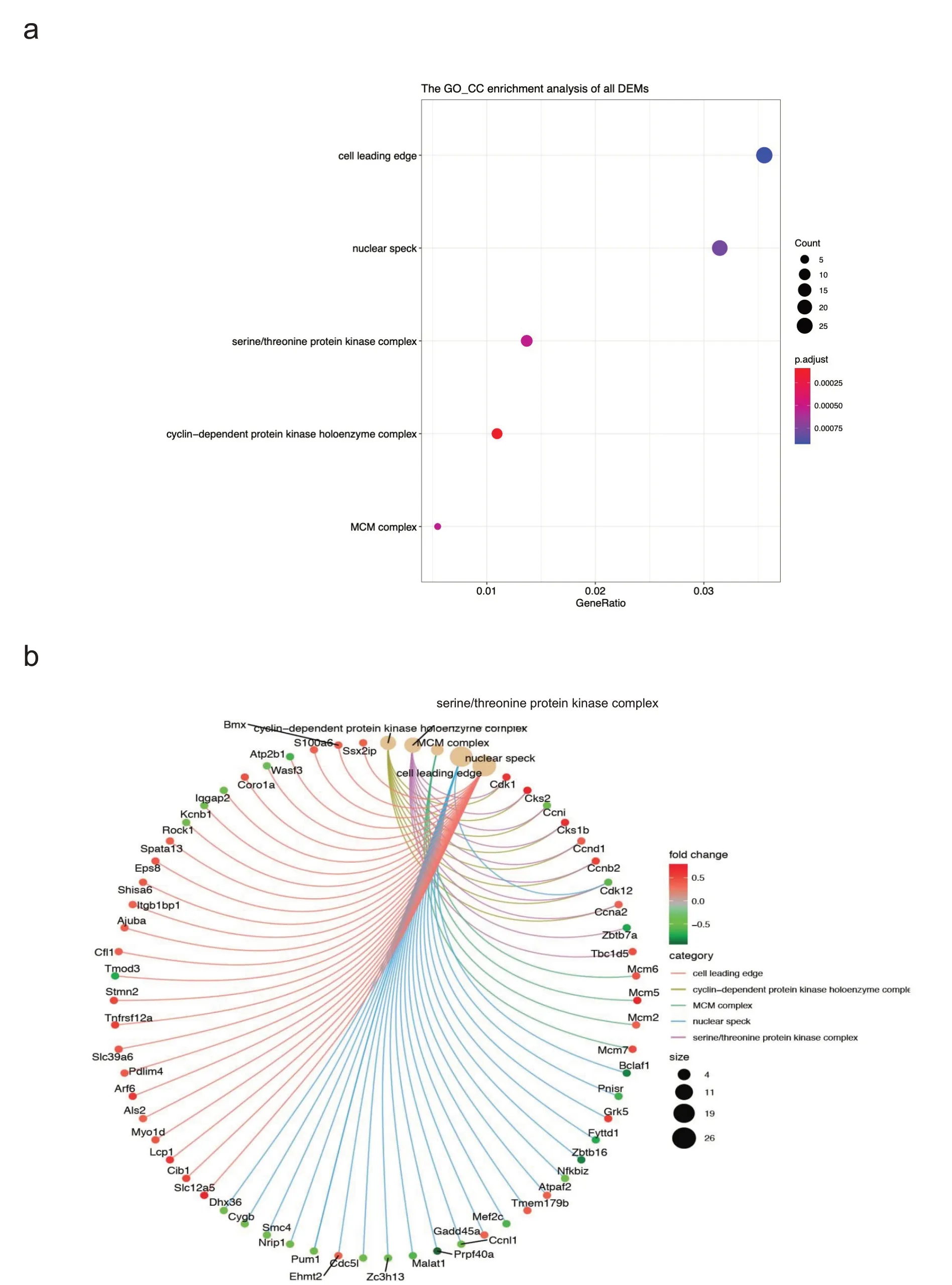
图4 自由转轮运动后小鼠心肌中差异基因的GO功能富集分析

图4 自由转轮运动后小鼠心肌中差异基因的GO功能富集分析(续)
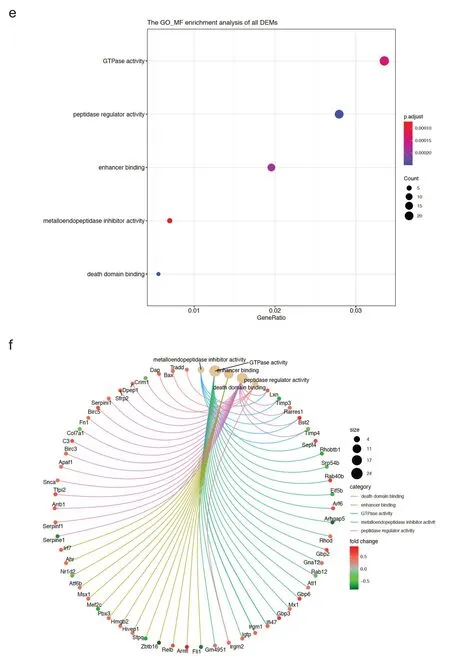
图4 自由转轮运动后小鼠心肌中差异基因的GO功能富集分析(续)
此外,KEGG 富集结果分析主要富集在细胞周期、补体和凝血级联、百日咳、p53信号通路和DNA复制等(如图5a 所示),差异基因与KEGG 富集结果的关系图如图5b所示。
2.5 游泳和自由转轮运动后小鼠心肌中差异基因的对比
为了观察游泳和自由转轮运动后小鼠心肌中差异表达基因的异同,本研究将游泳后上调基因(SM_Up)、游泳后下调基因(SM_Down)、自由转轮运动后上调基因(WR_Up)、自由转轮运动后下调基因(SM_Down)进行交互分析,分析结果如图6所示。结果显示,两种运动后小鼠心肌中共同上调的基因有8 个,分别是:Tmem176a、Snca、Zik1、Lgals3、C1qb、Atf6b、Cxcl10、Rom1;两种运动后小鼠心肌中共同下调的基因有10个,分 别 是:Pcf11、C77405、C78344、Klf9、Kdm5a、Crnkl1、Rock1、Dnajc3、Lgals4、Sp4;游泳后下调而自由转轮运动后小鼠心肌中上调的基因有7 个,分别是:Lst1、Slc39a6、Emr1、Ncs1、Psmb9、Inmt、Mx1。
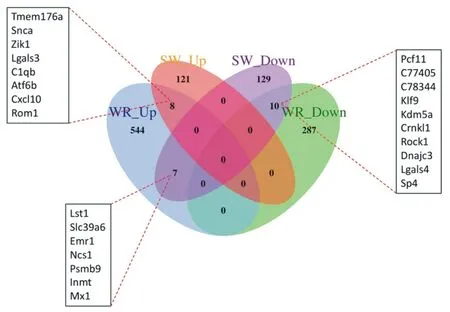
图6 差异基因韦恩图
2.6 差异基因网络互作分析
本研究分别对游泳和自由转轮运动后小鼠心肌中的差异基因进行了网络互作分析,并在R studio 中将结果可视化,如图7a 和7b 所示。在图中连线越多则节点越大,而连线越粗则互作得分越高,仅显示节点数大于5 的基因的名称。此外,在可视化过程中我们去除了单一互作关系和游离的互作关系。结果显示,游泳运动后有2 个节点数大于5 的基因,它们分别是:Agt和Pi3kr2;自由转轮运动后有7 个节点数大于5 的基因,它们分别是:Mad2l1、Spc25、Cdk1、Mcm6、Tipin、Cks2和Cd68。
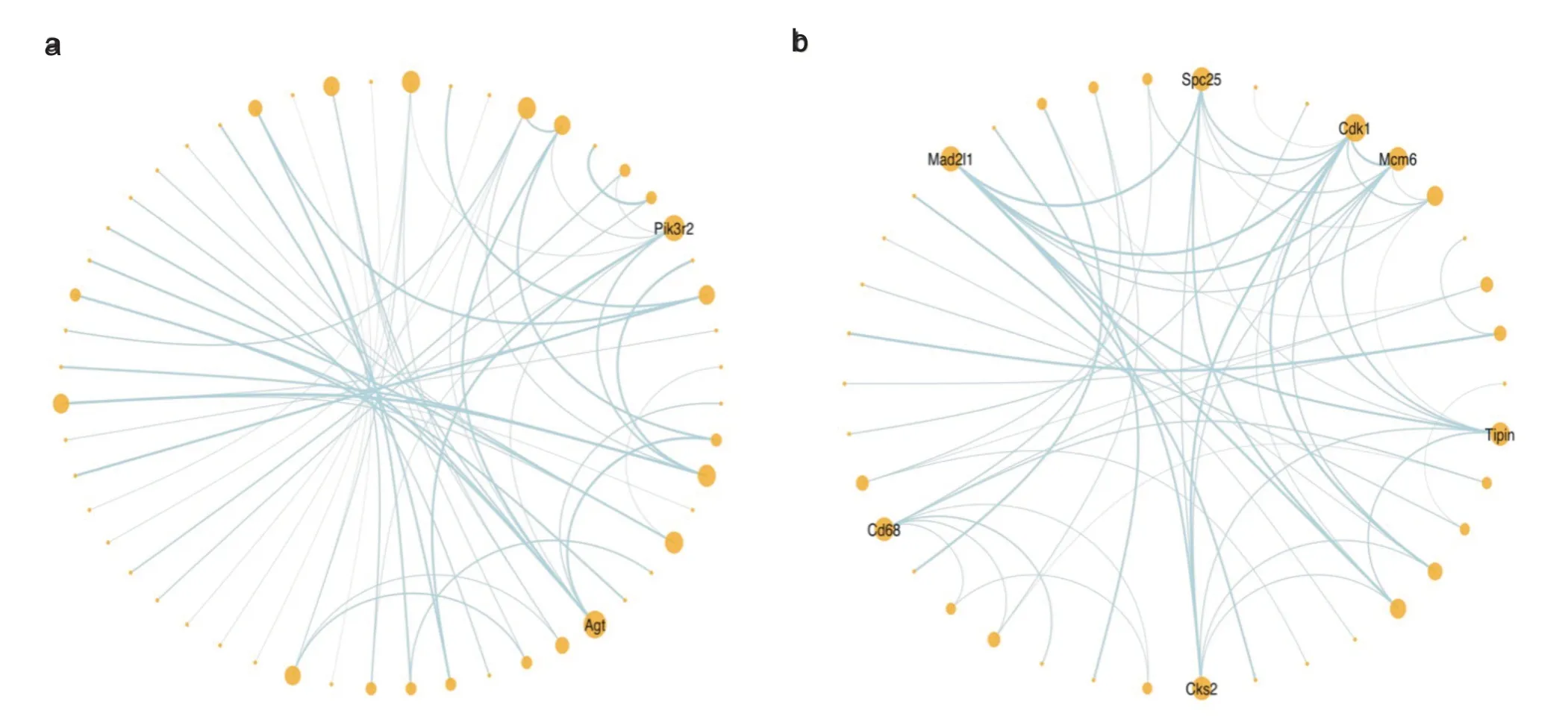
图7 差异基因网络互作图
2.7 差异基因的转录因子富集分析
最后本研究分别对游泳和自由转轮运动后小鼠心肌中的差异基因进行了转录因子富集分析,结果显示,游泳运动后小鼠心肌中的差异表达基因富集到的前3个转录因子分别是 predrem_nrMotif2039、predrem_nrMotif1919 和 dbcorrdb_TBP_ENOSR000EHA_1_m2,如图8a 所示;自由转轮运动后小鼠心肌中的差异表达基因富集到的前3 个转录因子分别是:jaspar_MA0050.2、homer_GAAAGTGAAAGT_IRF1 和taipale_cty_melth_IRF7_NNGAAANYGAAANYN_eDBD_m elth_repr,如图8b所示。
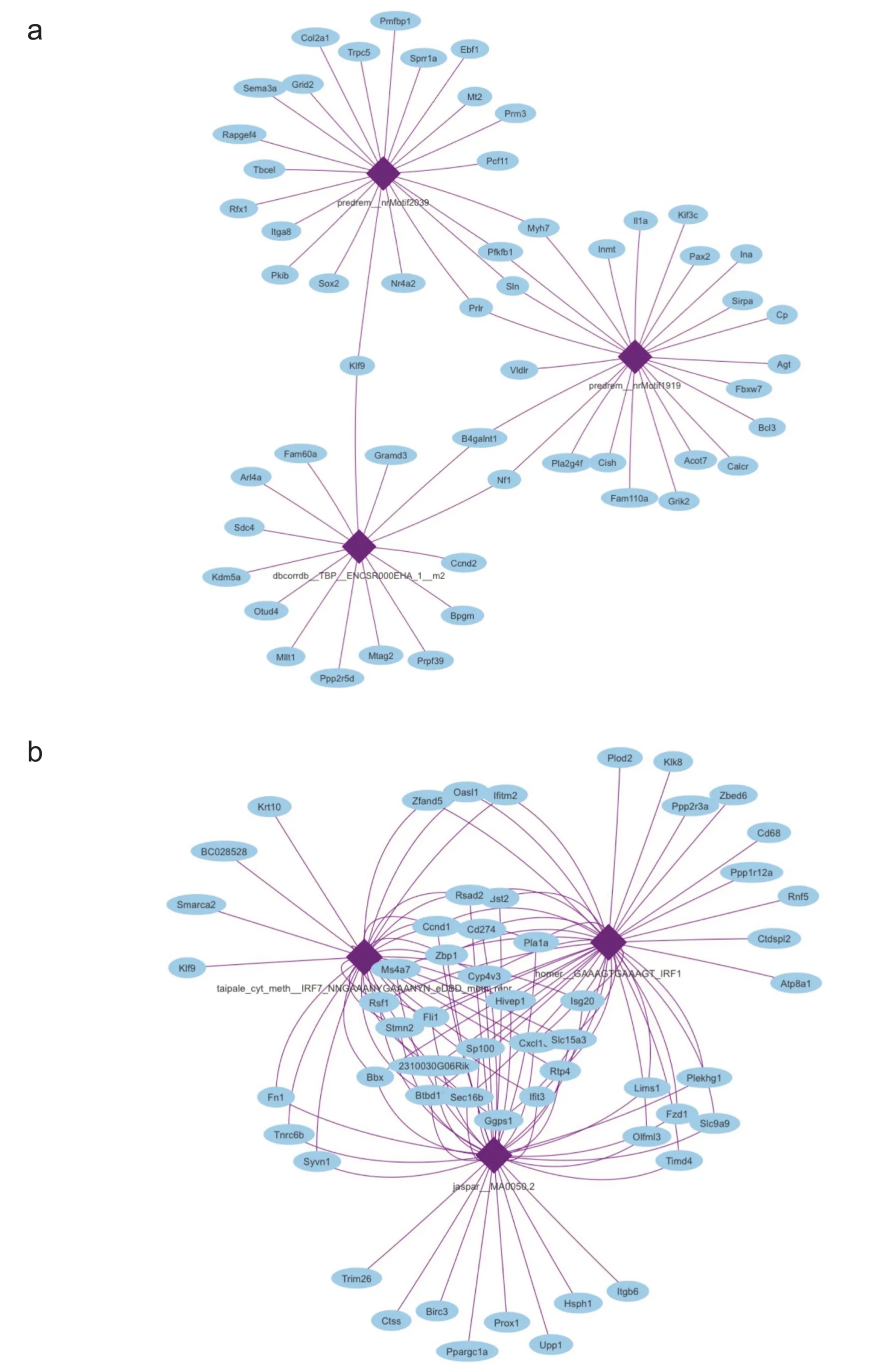
图8 转录因子富集分析
3 分析与讨论
本研究通过检索GEO 发现3 周游泳运动(GSE77)和3周自由转轮运动(GSE36330)的微阵列数据检测了成年雄性小鼠心肌中的基因表达情况,未发现3 周跑台运动后关于成年雄性小鼠心肌的微阵列数据。因此,本文对比了游泳和自由转轮这两种运动方式对运动心脏重塑过程中基因表达的影响。
通过对两个数据集的分析,本研究发现游泳运动后小鼠心肌中有275个差异表达基因,其中129个基因上调,146 个基因下调;而自由转轮运动后小鼠心肌中差异表达的基因有856个,其中559个基因上调、297个基因下调。从数量上来看,自由转轮运动后心肌中差异表达的基因多于游泳运动。随后本研究分别对这些差异基因进行了功能富集,从生物过程的富集情况来看,两者截然不同,游泳运动后小鼠心肌中的差异基因主要富集在血管生成和发育,而自由转轮运动后小鼠心肌中的差异基因主要富集在病毒反应。大量文献报道运动可促进心肌血管生成,本课题组前期研究也发现运动后心脏重塑过程中心肌毛细血管数量及毛细血管腔体积密度和表面积密度均出现了显着性增加[18]。本研究表明游泳运动后小鼠心肌中的差异表达基因可能参与了运动心脏重塑过程中血管生成和发育。然而,令人意外的是,自由转轮运动后心肌中差异表达基因的功能富集在了病毒反应,包括干扰素-b 和-g 反应,且本研究结果显示参与病毒反应的大部分基因的表达水平在自由转轮运动后升高,如图4d所示。尽管前期也有报道长期中强度有氧运动可增强机体免疫力,Zamani 等[19]也报道中强度运动会刺激外周血中的单核细胞生成更多的干扰素-g 和白介素-12(interleukin-12,IL-12),但目前没有文献报道运动心脏重塑过程中会伴随着病毒反应增强。因此,目前尚不清楚自由转轮运动诱导的心脏重塑过程中上调的这些病毒反应相关的基因及其调节的下游因子是否会释放到外周并随着循环作用于全身。此外,转轮运动增强心肌病毒反应的机制也尚不清楚。
为了进一步对比游泳运动和自由转轮运动对小鼠心肌基因表达的影响,本研究将两者的上调和下调基因进行了交互分析,结果显示两者共同上调的基因有8个,共同下调的基因有10个。考虑到两种运动方式都可以诱导生理性心肌重塑,本研究查看了两者共同上调的8个基因,然而在这8个基因中并未发现促进心肌细胞生长的基因,提示两种运动方式诱导的心脏重塑可能存在着不同的分子机制。因此,通过对不同运动方式诱导的心肌中差异基因取交集来获取运动心脏重塑的关键基因可能并不是一个理想的方法。
为进一步明确游泳运动和自由转轮运动在诱导心脏重塑过程中不同的分子机制,本研究分别对两者的差异表达基因进行了调控网络分析,并筛选出两者的关键基因。本研究发现游泳运动后心脏重塑的关键基因是血管紧张素原(angiotensinogen,Agt)和Pik3r2。Agt 最终可生成血管紧张素Ⅱ(angiotensin Ⅱ,AngⅡ),对于血管而言,AngⅡ具有很强的缩血管作用从而增加心脏的前负荷及后负荷。此外,文献报道AngⅡ还可直接作用于心肌细胞,并导致心肌重塑。AngⅡ通过与血管紧张素Ⅱ-1 型受体(angiotensinⅡtype 1 receptor,AT1R)结合激活G 蛋白耦联受体,进而导致蛋白合成和细胞生长。但是,过量的AngⅡ会导致心肌由生理性肥大转为病理性肥大,与此同时AT1R的表达水平也会显着升高[20]。本研究发现游泳运动后小鼠心肌中Agt表达水平升高,那么在运动过程中,AngⅡ/AT1R这一病理性信号通路将会发生怎样的变化?Li等[21]报道8周跑台运动后心肌中AngⅡ的表达水平显着升高,同时他们发现AT1R的数量及亲和力均显着下降,因而运动后高水平的AngⅡ并没有导致心肌病理性重塑。但是运动后心肌中Agt 水平升高并未导致病理性心肌重塑的具体机制还有待进一步的研究。
此外,本研究发现游泳运动诱导心脏重塑过程中的另一个关键基因Pik3r2(P85b)表达水平显着上升。Pik3r2 是PI3K 复合物的亚基之一,其表达水平上调后可负向调控PI3K/AKT信号通路,而这条通路被证实是心肌细胞生长的重要信号通路[14]。此外,Pik3ca(P110a)是PI3K 复合物的另一个亚基,它活化后可调控细胞增殖、分化和存活。本研究发现游泳运动后心肌中Pik3ca 的表达水平显着下降(图2b)。综合来看,本研究结果提示,3 周游泳运动后心肌中的PI3K/AKT信号通路可能是被抑制的。然而,McMullen 等[14]发现激活PI3K/AKT 信号通路是运动诱导生理性心肌肥厚的重要途径,敲除小鼠P110a基因后,尽管运动仍可引起心肌质量增加,但是其增加幅度不如野生小鼠大。Weeks等[22]发现PI3K/AKT信号通路对心肌具有保护作用,过表达P110a 基因可减轻动脉缩窄引起的心肌损伤,敲除P110a 基因则会加重动脉缩窄引起的心肌损伤。运动对动脉缩窄引起的心肌损伤也有保护作用,敲除P110a 基因后运动的心肌保护效应减弱。PI3K/AKT信号通路适度的激活对于心肌细胞的正常生长很有必要,然而其过度激活也会对心脏造成不利的影响。李奕等[23]报道PI3K蛋白水平在运动后上调然后回落,运动后24 h运动组PI3K蛋白水平与安静组无显着性差异,表明机体内存在一种机制在运动后抑制PI3K/AKT 信号通路防止其过度激活,最终实现运动后发生生理性不是病理性心肌肥大而。因此,本研究观察到游泳运动后Pik3ca基因的下调及Pik3r2基因的上调可能是运动后抑制PI3K/AKT信号通路的分子机制,这一解释尚待今后研究证实。
本研究发现在自由转轮运动后小鼠心肌重塑过程中共有7 个关键基因,分别是Mad2l1、Spc25、Cdk1、Mcm6、Tipin、Cks2 和Cd68。除了Cd68 外,其他6 个基因均参与细胞周期,表明自由转轮运动可能会促进心肌细胞的分裂和生长。可见,从差异基因网络互作结果上来看,游泳运动和自由转轮运动最终筛选出来的差异基因也是截然不同的。游泳运动可能是通过调节Agt 和PI3K/AKT 信号通路实现心肌细胞的生理性肥大,而自由转轮运动可能是通过促进心肌细胞的分裂和生长而实现心肌细胞的生理性肥大。
基因的表达受到转录因子的调控,为了进一步了解不同方式的运动对心脏重塑过程中基因表达变化的影响,本研究根据游泳和自由转轮运动后小鼠心肌中的差异基因分析它们所对应的转录因子。Kohli等[24]报道过一些影响心脏肥大的转录因子,包括促进心肌肥大的转录因子:GATA 家族(GATA-4/5/6)、MEF-2、Csx-Nkx-2.5、SRF /Myocardin、HAND、TEAD 和NFAT,以及抗心肌肥大转录因子:FoxO、MITF、YY1 和CHF1/Hey2。本研究结果显示游泳运动后小鼠心肌中的差异基因富集到的转录因子按AUC 排名前3 位的是predrem_nrMotif2039、predrem_nrMotif1919 和TBP,这三个转录因子在运动心脏重塑过程中的作用尚待进一步研究。自由转轮运动后小鼠心肌中的差异基因富集到的转录因子AUC排名前3位的是jaspar_MA0050.2、IRF1、IRF7,IRF 家族的转录因子主要参与免疫反应,这一结果与本研究差异基因功能富集到病毒反应相一致。
4 结论
通过对GEO 数据进行生物信息学分析,本研究发现游泳运动和自由转轮运动两种运动方式在诱导小鼠心脏重塑过程中差异基因的数量不同、共同差异基因少、差异基因功能富集及转录因子富集结果都截然不同,因此,这两种运动诱导的生理性心肌重塑的分子机制有可能也不同。



