荆婷婷,尹 号,黄 蓉,兰仕梅,李章程,尤留超,郝华芳,付 磊,储岳峰
(中国农业科学院兰州兽医研究所,兰州大学动物医学与生物安全学院,动物疫病防控全国重点实验室,兰州 730000)
牛支原体属于柔膜体纲、支原体属,是介于细菌和病毒之间的自然界存在的最小的最简单的能进行自我复制的原核微生物。目前已分离鉴定出与牛病相关的支原体有20多种,牛支原体是其中一种。1961年,美国人Hale首次从患乳腺炎的病牛中分离到牛支原体,1967年,发现其与牛呼吸道疾病有关,并被命名为Mycoplasmabovis[1],我国湖北省于2008年首次暴发了由牛支原体引起的犊牛“烂肺病”,发病率高达50%~100%,死亡率可达10%~50%,严重损害了我国养牛业的健康发展[2-3]。更多研究表明牛支原体能引起牛肺炎、乳房炎、关节炎、结膜炎等多种疾病,还可与其他病原一起引起混合感染[4-6],已给全球养牛业以及乳制品业造成了巨大经济损失[7-8],开展其致病机制研究及开发防控产品一直是科学工作者们研究牛支原体的热点方向。
牛支原体主要通过黏附宿主组织细胞来实现在机体中的定殖,进而破坏宿主免疫屏障。通过抑制宿主免疫、释放毒素物质、掠夺宿主营养物质等途经造成宿主损伤[9-13]。此外,在牛支原体膜表面存在着结构和功能的多样性可变脂蛋白家族(Vsps),有助于牛支原体躲避宿主免疫系统的识别。除了这些,牛支原体还存在一些毒力因子,如膜蛋白核酸酶,能诱导宿主细胞的凋亡[14]。早在1968年就有研究表明多种支原体具有溶血毒性,如肺炎支原体、鸡毒支原体、猪鼻支原体、绵羊肺炎支原体等[15-18]。孙远航等[18]原核表达了绵羊肺炎支原体Y98株的溶血素(hemolysin)HlyA基因,并证明了该蛋白具有溶血活性。猪鼻支原体通过同源重组定向缺失技术确定了与溶血相关的基因HlyC[20]。
本研究发现牛支原体模式菌株PG45以及08M、07801临床分离株对鼠红细胞有溶解能力,进而根据NCBI上的基因注释及序列比对找到假定的可能与溶血素相关的基因MBOVPG45_0580。通过对0580的氨基酸序列以及核苷酸序列进行分子特征分析,发现该蛋白氨基酸序列在牛支原体多种菌株中的保守性极高。通过原核表达外源性0580蛋白的C端截短体、制备其多克隆抗体、溶血试验、黏附试验等,初步分析MBOVPG45_0580在牛支原体致病机制中的生物学作用。
1 材料与方法
1.1 材 料
1.1.1 菌株、载体及实验动物 大肠杆菌感受态DH5α、BL21(AI),表达载体Pcold III,牛支原体PG45、08 M、07801菌株均来自本实验室。实验动物为6周龄BALB/c雌性小鼠,由中国农业科学院兰州兽医研究所动物实验中心提供。
1.1.2 主要试剂材料 氨苄青霉素(100 g·L-1)、IPTG诱导剂、咪唑、TBST、PBST、脱脂奶粉、牛血清蛋白(BSA)、LB培养基(胰蛋白胨10 g·L-1、酵母提取物5 g·L-1、氯化钠10 g·L-1)、TAE电泳缓冲液(2 mmol·L-1Tris、1 mol·L-1乙酸、100 mmol·L-1EDTA)、SDS电泳缓冲液(0.025 mol·L-1Tris、0.25 mol·L-1Glycine、0.5% SDS)、转膜缓冲液(48 mmol·L-1Tris、39 mmol·L-1Glycine、0.037%甲醇)等购自北京索莱宝科技有限公司;HigH Affinity NI-NTA Resin购自金斯瑞生物科技公司;蛋白质分子量标准、Super Signal West Atto超敏底物购自赛默飞尔而科技公司;HIS标签鼠源抗体、HRP标记的羊抗鼠抗体购自生工生物工程(上海)股份有限公司;DL 5000/2000 DNA marker、Phanta Super-Fidelity DNA聚合酶等试剂购自南京诺唯赞生物科技股份有限公司;质粒提取试剂盒、胶回收试剂盒、T4连接酶、NdeⅠ限制性内切酶、XbaⅠ限制性内切酶购自TaKaRa公司,CFDA SE细胞增殖与示踪检测试剂盒与DAPI核染料购自上海碧云天生物技术有限公司,鬼笔环肽染料购自美国Cytoskeleton公司。
1.1.3 主要仪器 CO2培养箱购自赛默飞生物科技有限公司,生物安全柜购自苏州安泰公司,PCR仪、垂直电泳系统、凝胶成像系统购自Bio-Rad公司,水平核酸电泳系统购自北京六一有限公司,激光共聚焦显微镜购自德国徕卡公司。
1.2 生物信息学分析以及同源性分析
以牛支原体PG45菌株基因组DNA为模板,从GenBank数据库查找PG45菌株0580蛋白的氨基酸序列(GenBank登录号:WP_013456244.1),将氨基酸序列上传至在线分析软件ProParam、ProScale analysis、SignalP 6.0,TMHMM Server 2.0,预测蛋白的理化性质、分子量大小、跨膜区等参数。利用DNAStar软件对0580蛋白的氨基酸序列、亲疏水区进行分析,筛选主要结构域。从NCBI数据库中收集支原体0580的氨基酸序列,采用MEGA 6.0软件进行氨基酸序列分析,并以此构建进化树。
1.3 引物设计以及重组蛋白0580-C原核表达载体的构建
将0580蛋白氨基酸序列交由生工生物工程(上海)股份有限公司按照大肠杆菌偏爱密码子进行密码子优化并进行基因合成,以便更容易在大肠杆菌中进行可溶性表达。通过Primer Premier 5.0软件设计扩增0580截短的C端基因的特异性引物,上游引物:0580-C-NdeI-F:5′AAACATATGGGCA-AGATCGGTCGTAATATTTACATCAC 3′;下游引物: 0580-C-His-XbaI-R:5′TTTTCTAGATCA-GTGGTGGTGGTGGTGGTGCTCGAGTTTATC 3′,预期扩增片段长为894 bp。PCR扩增0580-C片段,电泳后进行胶回收纯化。用限制性内切酶NdeⅠ、XbaⅠ分别对纯化产物和Pcold III载体进行双酶切,使用T4连接酶将酶切后的片段与载体过夜连接,转化到大肠杆菌感受态DH5α中,挑取菌落进行PCR鉴定,筛选出的阳性克隆进行测序。
1.4 重组蛋白0580-C的诱导表达与纯化
取50 ng测序完全正确的重组质粒转化入BL21(AI)表达菌株中,挑取单克隆于带有100 μg·mL-1氨苄青霉素抗性的3 mL LB培养基中进行复壮后转接,待菌液OD值到达0.6~0.8时,分别加入至终浓度为0、8、0.8、0.08 mmol·L-1的IPTG诱导剂,于16 ℃,120 r·min-1,诱导30 h。收集菌体,经超声破碎仪破碎后,制备上清液蛋白样和沉淀蛋白样。经过SDS-PAGE电泳染色后,观察蛋白是否表达,上清中蛋白表达量最多的则为最佳诱导剂浓度。使用最适表达条件进行大量诱导产生蛋白,采用NTA树脂进行亲和层析纯化。对纯化后蛋白进行超滤浓缩,使用Bradford法测浓缩后蛋白浓度检测浓缩后蛋白浓度。
1.5 重组蛋白0580-C的鉴定
将重组蛋白0580-C(rMbovP0580-C)以10 μL·孔-1上样,进行聚丙烯酰胺凝胶电泳,先用80 V电泳30 min再换成120 V继续跑胶1 h。通过湿式转印法将蛋白转印到PVDF膜上,电压为100 V,时间为1 h。将PVDF膜使用5% BSA在室温条件下封闭2 h后,使用商品化的鼠源一抗(HIS)按照1∶5 000稀释,室温孵育1 h,使用TBST洗涤3次,10 min一次。再用HRP标记的山羊抗鼠IgG二抗按照1∶5 000稀释,孵育1 h后洗涤,使用ECL进行显色。
1.6 多克隆抗体的制备与效价测定
在免疫前,对5只6周龄BALB/c雌性小鼠进行尾静脉采血,制备血清。首次免疫时,将纯化的蛋白与完全弗氏佐剂等体积充分混合形成油包水乳化剂,皮下多点注射进行免疫,免疫剂量为100 μg·只-1。之后第14、21、28、35天分别进行免疫,使用不完全弗氏佐剂与纯化的蛋白等体积混合,免疫剂量不变。最后两次免疫采用腹腔注射。第五次免疫一周后对小鼠进行眼眶静脉丛采血,分离血清,所得即为针对0580-C重组蛋白的鼠源多克隆抗体。
取12.5 μg纯化后的重组蛋白0580-C作为抗原进行包被,于4 ℃过夜孵育后,使用5%脱脂奶粉(0.137 mol·L-1NaCl,0.02 mol·L-1Tris,50 g·L-1脱脂奶粉)37 ℃封闭2 h。使用PBST洗涤3次,每孔300 μL,每次3 min。将获取的阳性以及阴性血清使用PBST从1∶800开始二倍比稀释进行孵育,每孔100 μL,37 ℃孵育1 h,洗涤后使用HRP标记的山羊抗鼠IgG(稀释倍数为1∶10 000)100 μL·孔-1再次孵育1 h。孵育结束后进行洗涤,使用显色液、终止液进行显色。最后用酶标测定仪在450 nm 处读取OD值。待测血清OD450 nm的数值记为S,阴性对照血清测定的OD450 nm数值记为N,S/N≥2.1即为阳性,以能获得阳性的最大稀释比例作为待检血清的抗体效价[21]。
1.7 重组蛋白0580-C的免疫印迹检测
将rMbovP0580-C以10 μL·孔-1上样,进行SDS电泳后转印到PVDF膜上,加入含有5% BSA的封闭液进行孵育;rMbovP0580-C的鼠源多克隆抗体按1∶3 000稀释后作为一抗,于摇床上(100 r·min-1)室温孵育1 h,使用TBST洗涤后再用HRP标记的山羊抗鼠IgG二抗按照1∶10 000稀释,孵育1 h后洗涤,使用ECL进行显色。
1.8 重组蛋白0580-C内源性表达鉴定
以1∶9的比例复苏牛支原体PG45、08 M、07801菌株,再转接至200 mL MTB培养基中,37 ℃培养72~84 h,集菌破碎后制备蛋白样,以10 μL·孔-1上样,进行SDS电泳,转印。加入含有5% BSA的封闭液进行孵育;将制备的rMbovP0580-C鼠源多抗按照1∶1 000稀释后作为一抗孵育1 h,洗涤后再用HRP标记的山羊抗鼠IgG二抗按照1∶10 000稀释,孵育1 h后洗涤,使用ECL进行显色。
1.9 重组蛋白0580-C的溶血性试验
将200、400 μg纯化超滤后的rMbovP0580-C分别与等体积的2%鼠红细胞悬液混合,于37 ℃温箱中孵育3 h,观察是否有溶血现象发生。
1.10 重组蛋白0580-C对PG45菌株黏附EBL细胞能力的影响
参考Zou等[22]、Zhu等[23]的研究方法设计蛋白对牛支原体黏附力影响的试验,并进行优化。
1.10.1 菌落计数法
1.10.1.1 支原体及细胞准备:将培养至对数生长期的牛支原体以12 000 r·min-1离心10 min,使用PBS洗涤3次后采用DMEM(无抗含血清)重悬,进行计数。将胎牛肺细胞(EBL)铺于12孔板中,待其生长至每孔约80%时,以DMEM(无抗无血清)洗涤3次并随机选取一孔使用胰酶消化计数。其余孔洗涤后加入细胞培养基。
1.10.1.2 细胞感染:感染量为每个细胞接种100个支原体,在阴性对照组加入400 μg BSA蛋白,试验组加入400 μg rMbovP0580-C,使用DMEM(无抗含血清)将每孔加液量补至1 mL,混匀后放到含5% CO2的37 ℃温箱中感染EBL细胞1.5 h。
1.10.1.3 计数:感染结束后,使用预热的PBS洗去未黏附的牛支原体,加入胰酶消化细胞,用细胞培养基终止消化并充分吹打细胞。取100 μL加入到900 μL PBS中,进行梯度稀释,取10-3、10-4稀释度悬液各100 μL涂布牛支原体固体培养基,置于37 ℃培养箱培养3~5 d后对菌落进行计数。
试验设置牛支原体PG45感染组(空白组)、PG45+BSA蛋白(阴性)组、PG45+ rMbovP0580-C(试验)组。每组设2个重复,试验平行重复3次,计算黏附率(%)=(试验组CFU÷输入PG45 CFU)×100%。
1.10.2 激光共聚焦显微镜观察法 按照CEDA SE试剂的说明书对牛支原体进行染色后,按照菌落计数法中所述进行细胞感染试验,其中rMbovP0580-C、BSA蛋白质用量为100 μg。感染结束后洗涤细胞,加入500 μL 4%多聚甲醛室温静置固定30 min。固定后加入300 μL 0.5% Triton 100,室温透化细胞10~15 min;洗涤后避光加入300 μL 鬼笔环肽对细胞骨架进行染色处理,室温反应20 min。洗涤后,避光加入300 μL DAPI细胞核染蓝色荧光染料,37 ℃反应20 min。洗涤干净后进行封片处理,于徕卡激光共聚焦显微镜下进行观察。
1.11 重组蛋白0580-C蛋白的多克隆抗体对PG45菌株黏附EBL细胞能力的影响
参考张懿[24]、Song等[25]试验方法设计并优化rMbovP0580-C的多抗对PG45黏附能力的影响试验。
1.11.1 菌落计数法 将针对rMbovP0580-C的鼠源多克隆抗体、鼠源阴性血清于56 ℃水浴锅中灭活30 min,将支原体与已灭活的血清按照1∶10的比例充分混合,放到含5% CO2的37 ℃温箱内共孵育1 h。向细胞中加入与血清共孵育后的PG45,使用DMEM(无抗含血清)将每孔加液量补至1 mL,于37 ℃温箱感染EBL细胞1.5 h。感染结束后对细胞进行洗涤、消化以及稀释涂板计数。
试验设置PG45(对照)组、PG45+免疫前血清(阴性)组和PG45+rMbovP0580-C多克隆抗体1∶10。每组设置2个重复,试验平行重复3次,计算黏附率(%) =(试验组CFU÷输入PG45 CFU)×100%。
1.11.2 激光共聚焦显微镜观察法 将支原体染色后与多抗血清按照1∶10的比例共孵育1 h,再感染EBL细胞1.5 h。按照“1.10”所述操作步骤对细胞进行固定、透化以及染色处理,于徕卡激光共聚焦显微镜下进行观察。
2 结 果
2.1 生物信息学分析以及同源性分析
PG45菌株的0580基因全长大小为1 293 bp,该基因编码430个氨基酸,预测的0580蛋白分子量约为49 ku,等电点为5.99,不稳定系数为39.03,是一种稳定的亲水性蛋白(图1A)。经TMHMM Server v2.0、SMART Model和SignalP 6.0 Sever预测,该蛋白含有4个跨膜区,N端的1—28氨基酸序列预测为信号肽区域(图1B),C端蛋白截短体含有289个氨基酸,大小约为37 ku,存在两个CBS结构域(图1C),CBS编码胱硫醚-β-合成酶,参与同型半胱氨酸的转硫途经。通过Blastp将0580蛋白序列比对nr数据库,用MEGA 5.0软件进行序列比对分析并构建进化树(图1D)。如图所示,0580蛋白氨基酸序列非常保守,在08M、CQ-W70、HB-0801、PG45这些菌株中0580相似性为100%;同样与无乳支原体、与猪鼻支原体、绵羊肺炎支原体、猪肺炎支原体相似性较高。
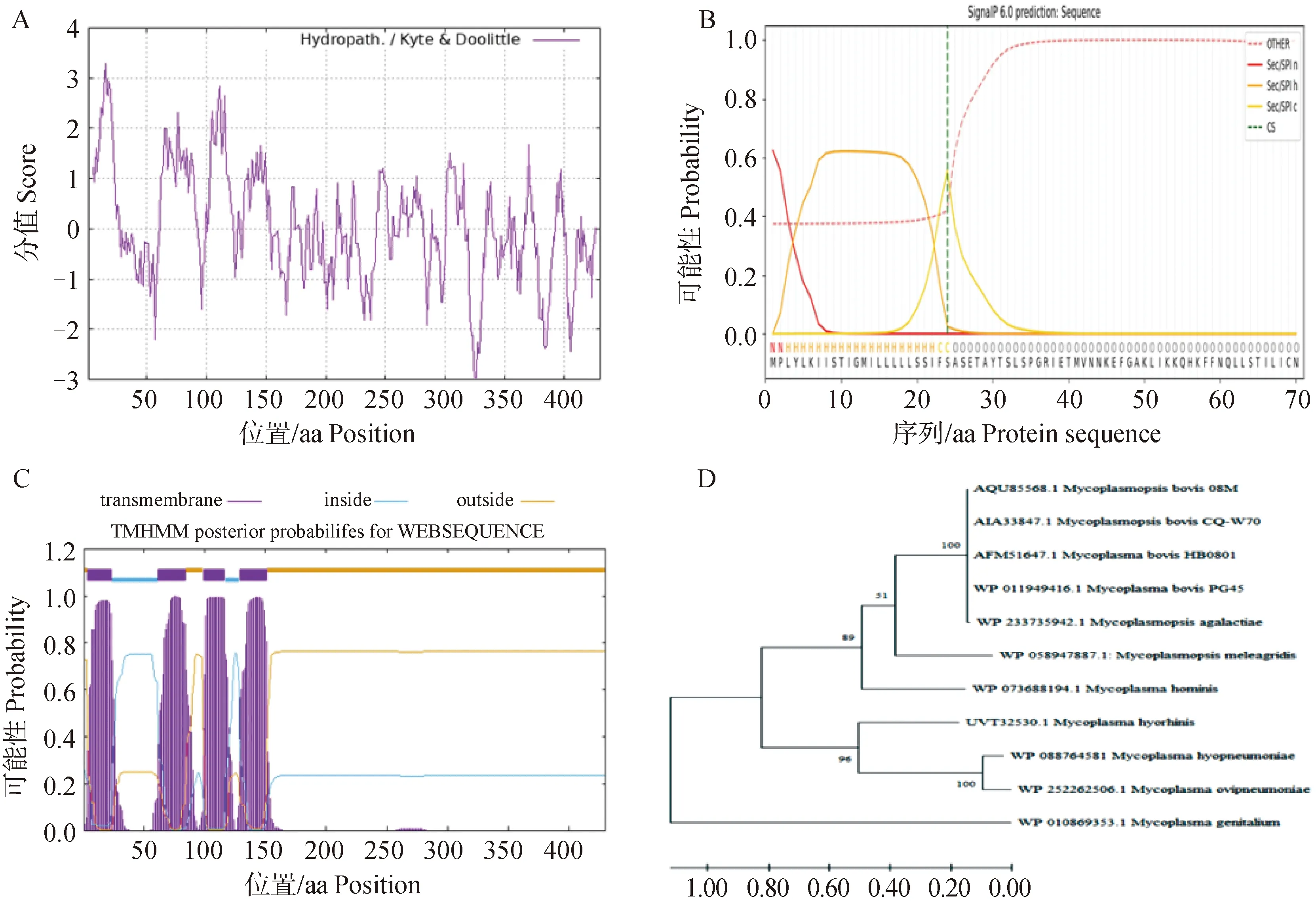
A. 0580蛋白亲/疏水性预测;B. 0580蛋白信号肽预测;C. 0580蛋白跨膜结构预测;D. 基于0580氨基酸序列构建进化树A. Hydrophilicity and hydrophobicity prediction of 0580 protein; B. Signal peptide prediction of 0580 protein; C. Transmembrane domain prediction of 0580 protein; D. Phylogenetic tree based on 0580 amino acid sequences图1 rMbovP0580-C生物信息学分析Fig.1 Bioinformatics analysis of the 0580 protein
2.2 rMbovP0580-C原核表达载体构建
使用引物对0580-C基因进行PCR扩增,片段大小为894 bp(图2A),符合预期大小。将0580-C基因片段连接到Pcold III载体上,通过PCR鉴定与测序比对,成功构建0580-C原核表达载体(图2B)。
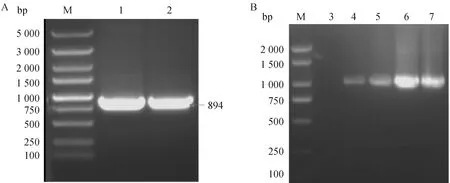
A. 0580-C基因片段扩增;B. PCR鉴定单菌落;M. DL5000 DNA相对分子质量标准;1~2. 0580-C基因扩增片段;3. H2O;4~7. 阳性转化子A. PCR amplification result of 0580-C gene; B. Identified the clony by PCR; M. DL5000 DNA marker; 1-2. 0580-C gene; 3. H2O; 4-7. Positive transformation 图2 0580-C基因扩增及重组质粒的构建Fig.2 Amplification of 0580-C gene and construction of recombinant plasmid
2.3 rMbovP0580-C的诱导表达与纯化
经SDS电泳分析,rMbovP0580-C在IPTG浓度为8、0.8、0.08 mmol·L-1时均进行表达(图3A)。选取0.8 mmol·L-1IPTG进行诱导表达、纯化,蛋白质相对分子质量约为37 ku,符合预测值(图3B)。
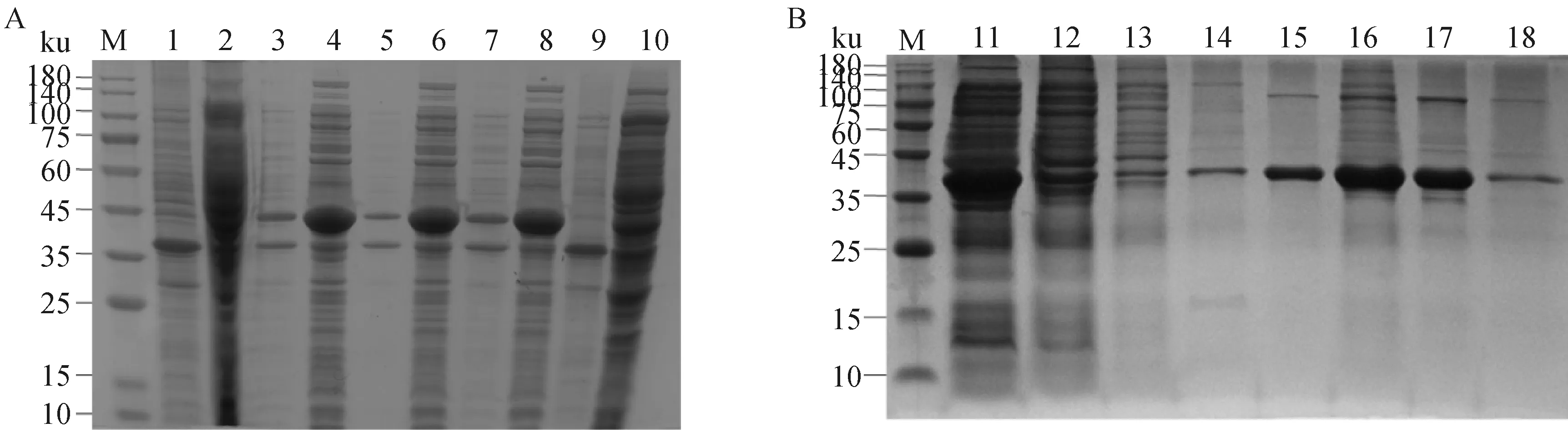
A. 在不同IPTG浓度下rMbovP0580-C的表达;B. rMbovP0580-C的纯化;M. 蛋白质相对分子质量标准; 1、3、5、7. IPTG浓度分别为0、8、0.8、0.08 mmol·L-1时的诱导表达破碎后的上清蛋白;2、4、6、8. IPTG浓度分别为0、8、0.8、0.08 mmol·L-1时的诱导表达破碎后的沉淀;9~10.空载质粒对照; 11.菌体裂解后上清液;12.过柱液;13~18.咪唑浓度分别为12.5、50、100、200、300、1 000 mmol·L-1的洗脱液A. Expression of recombinant 0580-C protein under different concentrations of IPTG; B. purified the recombinant 0580-C protein;M. Protein marker; 1, 3, 5, 7. Supernatant of protein was induced by 0, 8, 0.8, 0.08 mmol·L-1 IPTG; 2, 4, 6, 8. Precipitation of protein was induced by 0, 8, 0.8, 0.08 mmol·L-1 IPTG; 9-10.Control; 11. Supernatant; 12. Effluent; 13-18. Eluent of 12.5, 50, 100, 200, 300, 1 000 mmol·L-1 imidazole图3 rMbovP0580-C的表达与纯化Fig.3 Expression and purification of rMbovP0580-C
2.4 rMbovP0580-C的外源性及内源性表达鉴定、免疫印迹检测与效价测定
以小鼠抗His单抗为一抗、HRP标记山羊抗小鼠IgG抗体为二抗,进行SDS PAGE及Western blot分析,在37 ku附近产生特异性反应条带(图4A、B),成功表达重组蛋白0580-C。
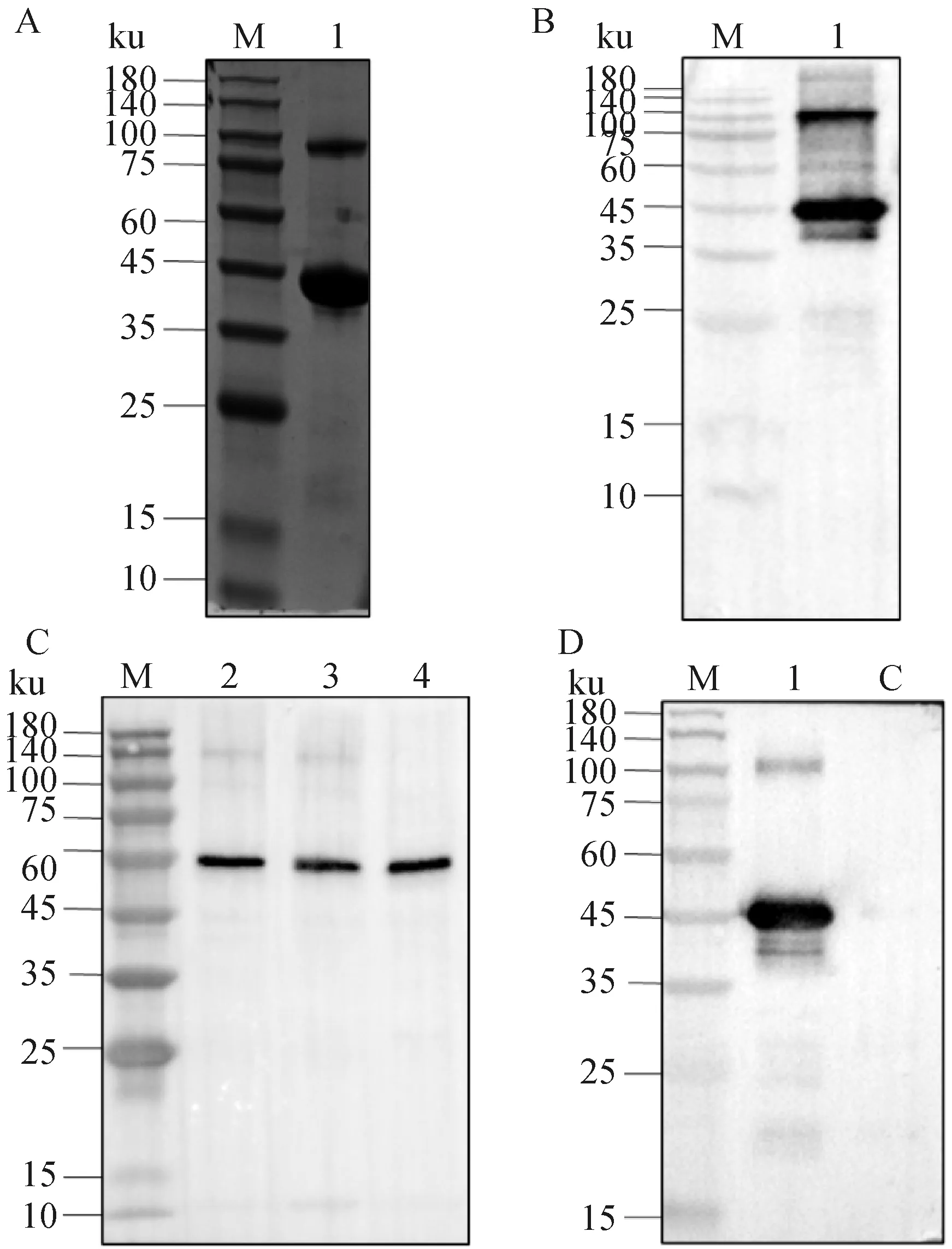
A. SDS PAGE检测rMbovP0580-C蛋白的表达;B. Western blot检测rMbovP0580-C蛋白的表达;C. rMbovP0580-C多克隆抗体检测内源性表达表达的0580蛋白;D. rMbovP0580-C多克隆抗体检测重组表达的rMbovP0580-C蛋白;M. 蛋白分子量标准;1. 纯化的rMbovP0580-C蛋白;2~4. 08M、07801、PG45上清液; C.对照蛋白A. Detection the expression of rMbovP0580-C protein by SDS PAGE; B. Detection the expression of rMbovP0580-C by Western blot; C. rMbovP0580-C polyclonal antibody was used to detect the endogenous expression of rMbovP0580-C protein; D. rMbovP0580-C polyclonal antibody was used to detect the recombinant rMbovP0580-C protein; M. Protein marker; 1. rMbovP0580-C purified protein; 2-4. Supernatant of 08M, 07801, PG45; C. Control protein图4 rMbovP0580-C 蛋白的鉴定与多克隆抗体的特异性检测Fig.4 Identification of rMbovP0580-C protein and detection of specificity of polyclonal antibody
为检测针对rMbovP0580-C产生的多克隆抗体的特异性,使用多抗检测牛支原体不同菌株。Western blot结果显示:鼠源 rMbovP0580-C多克隆抗体能够检测出牛支原体PG45、08M、07801菌株中的天然0580蛋白,条带在60 ku附近,大于预测大小(图4C)。
将制备的鼠源rMbovP0580-C的多克隆抗体与rMbovP0580-C进行Western blot反应,出现特异性条带(图4D)。
取12.5 μg rMbovP0580-C蛋白作为抗原包被,鼠源rMbovP0580-C多克隆抗体血清作为抗体,进行间接免疫ELISA试验,结果显示:鼠源rMbovP0580-C的多克隆抗体效价达1∶204 800(图5A),表明rMbovP0580-C蛋白免疫原性良好。
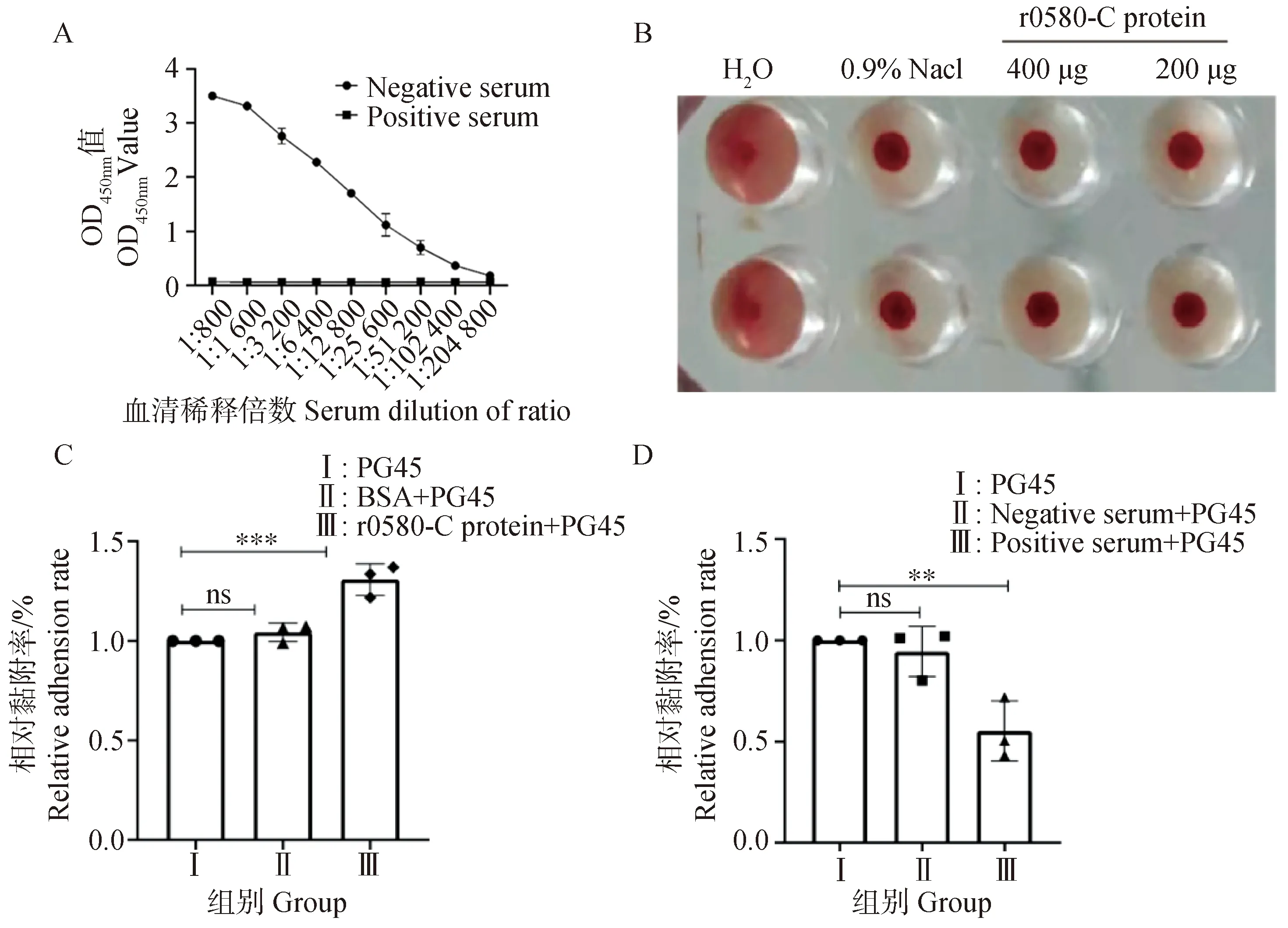
A. rMbovP0580-C多克隆抗体效价测定;B.鼠红细胞溶血性试验;C. rMbovP0580-C蛋白对PG45菌株黏附EBL能力的影响;D. rMbovP0580-C多克隆抗体对PG45菌株黏附EBL能力的影响A. Titer detection of rMbovP0580-Cpolyclonal antibody; B. Hemolytic test of mouse red blood cells; C. Effect of rMbovP0580-C protein on PG45 strain adhesion of EBL cell; D. Effect of rMbovP0580-C polyclonal antibody on PG45 strain adhesion of EBL cell图5 rMbovP0580-C多抗效价的测定及rMbovP0580-C蛋白生物学功能初步分析Fig.5 Titer detection of rMbovP0580-Cpolyclonal antibody and preliminarily analyse biological function of rMbovP0580-C protein
2.5 rMbovP0580-C生物学功能初步分析
将0580-C蛋白与鼠红细胞37 ℃共孵育后发现,rMbovP0580-C蛋白不具有溶解鼠红细胞的能力(图5B)。但是通过分析细胞黏附及黏附阻断试验的结果发现,加入 rMbovP0580-C蛋白后,牛支原体PG45菌株对EBL细胞的黏附作用增强(图5C);而加入 rMbovP0580-C抗血清后,PG45菌株对EBL细胞的黏附能力下降(图5D)。激光共聚焦显微镜观察结果显示,当加入rMbovP0580-C重组蛋白后,细胞周围的绿色荧光数量增多,即黏附的支原体增多;而将牛支原体与rMbovP0580-C的多抗血清孵育后再感染细胞,细胞周围的绿色荧光减少,即多抗血清对牛支原体对EBL细胞的黏附有抑制作用(图6B)。
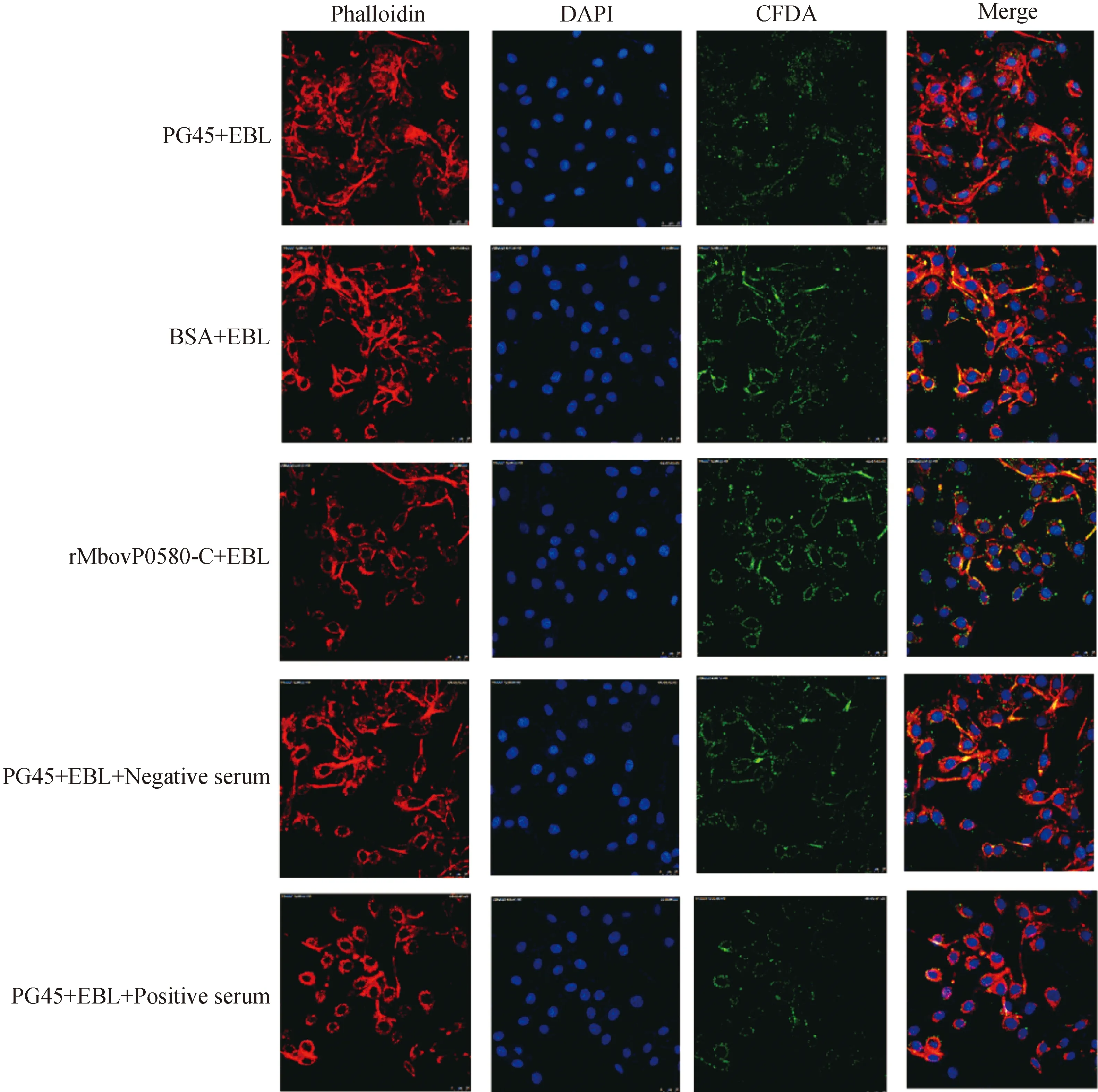
图6 rMbovP0580-C细胞黏附及黏附阻断试验Fig.6 Cell adhesion and blocking of rMbovP0580-C
3 讨 论
本实验室发现牛支原体能够使鼠红细胞溶血,通过基因注释找到了假定的与溶血素相关的基因序列MBOVPG45_0580,尝试对其进行全长表达。由于该基因编码的蛋白具有4个跨膜区,在温度、诱导剂浓度、摇床振荡速度等条件的优化下,更换Pcold载体、PET28a载体、PGEX载体均无法表达0580基因编码的全长蛋白。作者对该蛋白结构进行分析,发现在其C端具有两个CBS结构域,CBS (cystathionine-β-synthase)是编码胱硫醚-β-合成酶的基因,参与半胱氨酸的代谢途经。绵羊肺炎支原体溶血素TlyC基因也含有CBS结构域[26],在加入半胱氨酸时,穿透支原体对羊红细胞溶血作用提高了10倍,表明穿透支原体的溶血素可能含有半胱氨酸残基[16]。因而作者对含有CBS区的0580-C端截短体进行原核表达,纯化后的 rMbovP0580-C蛋白免疫小鼠后能得到高效价的抗体,免疫原性良好,但截短后外源表达的rMbovP0580-C并不具有溶血功能。黏附是牛支原体侵袭宿主组织的第一步[27],目前,已报道的与牛支原体黏附作用有关的蛋白有16种[28],如α-enolase[25]、TrmFO[29]、FBA[30]、P27[32]、P48[32]、NADH[33]和NOX2[34]等。本研究中的细胞黏附以及黏附阻断试验结果显示:0580-C蛋白可促进牛支原体对EBL细胞的黏附,而其多克隆抗体有黏附抑制作用,故推测0580基因编码的蛋白可能在PG45对宿主的黏附过程中发挥促进作用,进而助于感染宿主细胞。在本实验室构建的牛支原体突变体库中未筛选到假定的与溶血素相关的突变株,因此无法对该基因进行深层次的机制探究。基于牛支原体的定点突变的遗传操作工具的缺乏,制约了人们对牛支原体基因功能的探索,希冀学者在此方面有更进一步的研究。
4 结 论
成功地原核表达出牛支原体0580基因编码的假定与溶血素相关的蛋白C端截短体,尽管rMbovP0580-C蛋白不能使鼠红细胞溶血,但能促进PG45菌株对胎牛肺细胞的黏附能力,表明rMbovP0580-C蛋白可能是牛支原体的一种黏附分子,这为深入研究假定溶血素蛋白的生物学功能及牛支原体致病机制提供了研究基础。



