邵 静,张 颖,唐 毓,许保增
(中国农业科学院特产研究所,长春 130112)
卵母细胞是现阶段所能观察到的雌性哺乳动物体内最大的细胞,也是唯一的生殖细胞。和体细胞相比,卵母细胞的数量非常有限,并且不会再增殖。随着体外受精、动物克隆与转基因动物生产等生物学技术的快速发展,卵母细胞的重要性被体现的淋漓尽致。一颗获得良好发育的成熟卵母细胞是保障受精的顺利进行、双倍体合子的形成和早期胚胎基因组激活的物质前提[1]。卵母细胞的成熟受到一系列机制细致精密的调控,是多种信号通路相互协同又互相拮抗的结果。在受精过程中,只有细胞核与细胞质均发育成熟的卵母细胞才会对精子的结合释放出正确的应答反应,共同形成一个具有发育潜能的受精卵[2]。
很多研究认为,ERM蛋白参与了卵母细胞成熟和受精的过程。ERM家族是埃兹蛋白(ezrin)、根蛋白(radixin)和膜突蛋白(moesin)的统称,几乎所有的哺乳动物中都存在ERM蛋白,并且这3个蛋白的序列高度保守,共有序列高达75%,使它们在功能上非常相似[3]。ERM蛋白凭借其特殊的分子结构,可以直接或间接地与跨膜蛋白和肌动蛋白结合,从而作为“脚手架”,形成膜蛋白-ERM蛋白-细胞骨架蛋白复合体,参与构建并维持细胞皮层。细胞皮层通常被定义为质膜下方比较致密的肌动蛋白细胞骨架层,由微丝和微丝结合蛋白交联成凝胶状态的三维网状结构,在质膜与胞质之间形成了均匀的细胞质皮质区[4]。细胞皮层可以参与细胞形态的改变、细胞间的相互作用以及细胞膜对细胞的调节作用。ERM作为细胞皮层的核心组分,广泛分布在细胞质基质、片状伪足、丝状伪足和微绒毛中,控制肌动蛋白细胞骨架动力学,参与细胞膜的组成、囊泡形成、细胞迁移、信号传递、跨膜蛋白定位和皮质张力调节等生理过程[5]。
哺乳动物卵母细胞有高度组织化的细胞皮层,包括密集的微绒毛排列和复杂的微丝帽结构。在哺乳动物卵母细胞成熟过程中,纺锤体要靠皮质肌动蛋白的牵拉远离细胞中心,实现胞质不对称减数分裂[6]。在哺乳动物受精过程中,卵母细胞质膜和细胞骨架之间的关联通路可以保障精子与卵子的成功融合,例如,精子需要与卵质膜上的微绒毛结合才能进入卵母细胞[7],而微绒毛和细胞内部的信号传递,需要ERM蛋白将质膜、跨膜蛋白和基底的肌动蛋白串联起来,形成完整的调节路径,改变膜蛋白的动力性能[8]。这类生殖活动普遍都有ERM的参与,但是ERM蛋白家族在其中发挥的具体作用机制却很少受到深入研究。本文将以ERM蛋白为中心,概述其在哺乳动物卵母细胞减数分裂成熟和受精过程中参与的各种调控作用。
1 ERM蛋白的分子结构及特点
1.1 ERM蛋白的分子结构
ERM蛋白能够广泛参与细胞的多个生理过程都是依赖于它特殊的组成结构。在人类中,ezrin、radixin和moesin蛋白分别由585、583和577个氨基酸残基组成,ERM蛋白质家族的氨基(N)末端是由大约300个氨基酸残基组成的球状FERM结构域,该结构域由3个子域(F1、F2和F3)一起交叠成三叶草的形状;羧基(C)末端是由大约100个氨基酸残基组成的C-ERMAD结构域;两端中间是约150个氨基酸残基组成的α-螺旋结构,不同物种间ERM蛋白的序列差异几乎都来自于这段区域[9]。
ERM有两种构象状态:一种非活性状态,大多数细胞的ERM蛋白都是这种休眠状态,其N端的FERM结构域和C端的C-ERMAD相互抑制,以头对尾的方式结合起来,α螺旋区盘绕在它们中间,掩盖了它们与细胞骨架和细胞质膜的结合位点,成为闭合构象的失活状态,在细胞质中以单体环或二聚体的形式存在;另一种是定位在质膜上的非折叠的活性状态,其激活的标志是ezrin、radixin和moesin中第567、564、558位的苏氨酸残基被磷酸化,C-ERMAD从FERM结构域中释放出来,暴露出两端的结合位点[10](图1)。
1.2 ERM蛋白介导质膜与肌动蛋白的连接
肌动蛋白是细胞骨架的主要成分,多以球形单体(G-肌动蛋白)和聚合纤维(F-肌动蛋白)两种形式存在于真核生物中,肌动蛋白聚合成微丝,在细胞表面形成应力纤维、板状伪足、丝状伪足、黏着斑、微绒毛、分裂环等结构,参与细胞分裂和细胞迁移等许多生物学过程[12],卵母细胞的减数分裂和受精也离不开复杂的微丝网络。
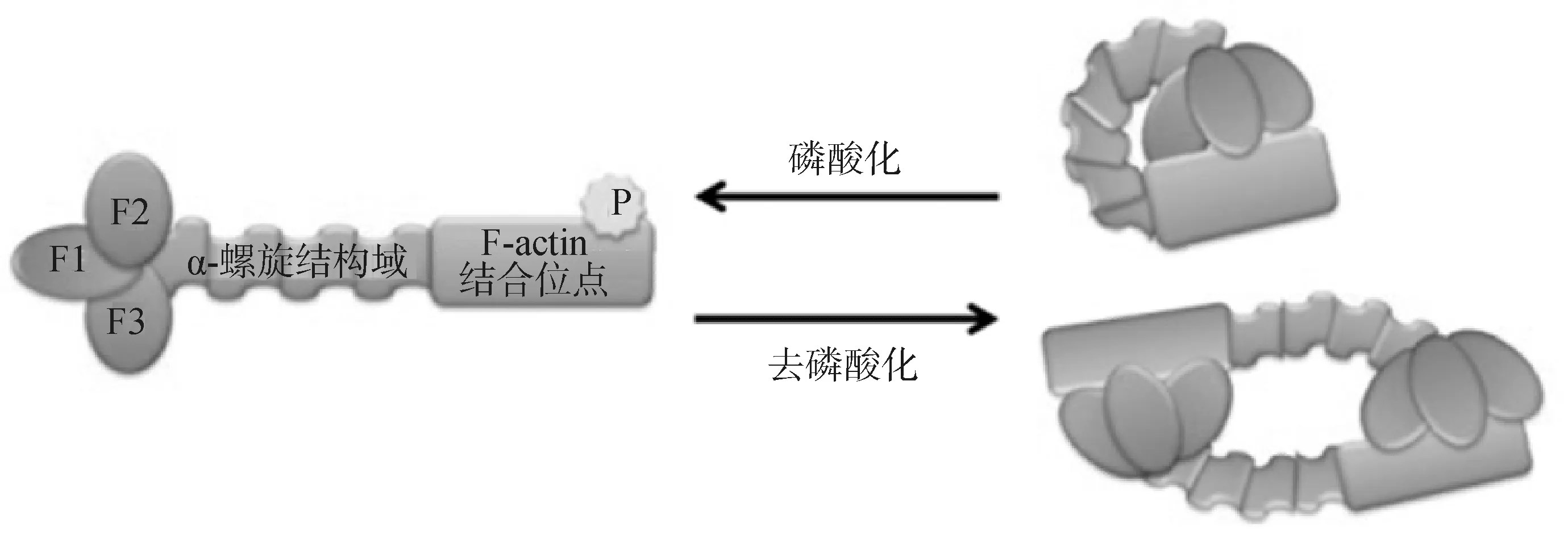
图1 ERM蛋白的两种构像示意图[11]Fig.1 Diagrams of two conformations of ERM proteins[11]
ERM蛋白在磷酸化与非磷酸化状态中的可逆转变,是调节质膜与肌动蛋白相互作用的关键。能够控制ERM活性的分子有很多,包括与肌动蛋白组装有关的Ran GTPase[13],与细胞极化有关的趋化因子,以及针对苏氨酸的磷酸酶、激酶和它们的抑制剂[14]。当ERM蛋白C-ERMAD结构域的苏氨酸被去磷酸化,ERM会从细胞皮层脱离,游离于细胞质中。“开放”的ERM蛋白FERM结构域与酸性的磷脂酰肌醇-4,5-二磷酸(PI(4,5)P2)结合并经过修饰后,可以促进C末端保守的苏氨酸残基发生磷酸化,磷酸化的C-ERMAD结构域带上负电,与FERM区域的负电荷发生排斥,使ERM蛋白的折叠构象打开[15]。
ERM蛋白C末端结构域包含肌动蛋白结合位点,而ERM的N末端FERM结构域可以与跨膜蛋白留在细胞质部分的带正电荷氨基酸簇的尾部直接结合,也可以通过先与支架蛋白EBP50或E3K-ARP结合,然后利用该蛋白的PDZ结构域间接关联到膜蛋白[16]。与ERM结合的膜蛋白很多都是粘附分子,包括CD44、CD43、PSGL-1和ICAM-1-3等[17-19]。目前的研究表明,ERM蛋白是连接整合膜蛋白与作为细胞骨架的皮质层肌动蛋白的接头蛋白。许多跨膜蛋白都是通过与胞质中的细胞骨架蛋白结合来改变细胞形状,并参与细胞粘附、细胞运动并维持细胞表面的信息交流,包括胞吞和胞吐作用[20-21]。ERM蛋白通过同时与整合膜蛋白、膜相关细胞质蛋白和肌动蛋白丝之间相互结合,组建专门的膜结构域,将胞外信号输送到细胞内,从而介导细胞迁移[22]、细胞分裂[23]、信号传导[24]、跨内皮细胞迁移[25]、微绒毛形成[26]以及细胞极性的建立[27]等重大生物学事件。
2 ERM蛋白参与卵母细胞成熟
卵母细胞的成熟指的是处于第一次减数分裂前期的双线期,也就是生发泡期(GV)的卵母细胞经历连续两次减数分裂并最终停滞在减数第二次分裂中期(MⅡ)的过程(图2)。
2.1 ERM蛋白调控卵母细胞-颗粒细胞间的双向信号传导
哺乳动物从出生就已经具备了一生的原始卵泡池,里面含有处于GV期的休眠卵母细胞和包裹着它的尚未分化的前颗粒细胞。原始卵泡是雌性哺乳动物的基本生殖单位[29]。卵母细胞只有接收到周围颗粒细胞释放的调节信号,才能突破休眠开始发育。该过程包含复杂的信号通路。前颗粒细胞受到营养、压力、能量、生长因子等信号刺激后,激活mTORC1信号通路,进而分化为颗粒细胞[30]。mTORC1信号通路继续促进颗粒细胞分泌生长因子KIT配体,与休眠卵母细胞表面的KIT受体结合,激活卵母细胞内的PI3K信号通路,下游FOXO3a被磷酸化并从卵母细胞的细胞核中转移到细胞质中,使休眠的卵母细胞开始生长,原始卵泡被激活为初级卵泡[31-32]。此时的颗粒细胞开始表达缝隙连接蛋白43(CX43),将彼此相邻的颗粒细胞连通起来;同时卵母细胞表达CX37蛋白,形成卵母细胞-颗粒细胞间的缝隙连接[33]。磷酸化的ezrin蛋白可以与CX43结合,而中间的α-螺旋区有一个AKB结构域,可以与蛋白激酶A(PKA)的调节亚基结合,ezrin蛋白将PKA锚定在下游,形成PKA-ezrin-CX43复合体,cAMP激活PKA后,使PKA释放出两个催化亚基将CX43在369和373位的丝氨酸残基磷酸化[34-35]。ezrin蛋白就以这种方式参与由cAMP/PKA信号通路介导的缝隙连接。在缝隙连接的介导下,颗粒细胞将能够调节卵母细胞pH和渗透压的离子、氨基酸和葡萄糖等分子以及保持卵母细胞长时间停滞在GV期所需的cGMP和cAMP传递到卵母细胞中[36]。但是透明带的形成使卵母细胞-颗粒细胞间有了一层物理性屏障,为了克服这层阻碍,透明带中出现了两种不同来源的突起状结构(图3)[37]。一种是由颗粒细胞发射出的跨透明带的丝状伪足(granulosa cell transzonal projections, GC-TZPs);另一种则是卵母细胞产生的状似蘑菇的顶端带囊泡的微绒毛结构(oocyte microvilli, Oo-Mvi)[38]。与颗粒细胞TZP相比,Oo-Mvi要稀疏很多。
微绒毛是细胞质伸出的微细指状突起,表面被细胞膜包裹,内部为细胞质,轴心是肌动蛋白聚合形成的微丝,广泛存在于动物细胞的表面,在卵母细胞和上皮细胞等吸收代谢机能旺盛的细胞中尤其常见[39]。因为微绒毛的存在可以增加细胞的表面积,同时微绒毛也可以伸缩和摆动,使细胞与外界微环境充分接触,所以富集微绒毛的细胞一般都需要频繁进行细胞与细胞间的物质交换和信息交流。附着在卵母细胞表面的微绒毛比体细胞还有一系列特殊的细胞行为。
微绒毛在卵母细胞不同发育阶段所起的主要作用是不同的。未成熟的卵母细胞会通过微绒毛从周围的颗粒细胞中获取营养物质和信息分子以满足自身的发育需求,也会通过微绒毛释放生长分化因子9(GDF9)和骨形态发生蛋白15(BMP15)等分泌因子,促进卵泡生长和颗粒细胞增殖[40]。如果阻断颗粒细胞对卵母细胞的联系,微绒毛就会退化,颗粒细胞提前黄体化,这颗卵泡将来会成为不能排卵的闭锁卵泡[41]。排卵前卵母细胞表面的微绒毛会塌陷回缩,从而切断与卵丘细胞间的联系,方便其从卵泡中排出[42]。排卵后2~4 h,卵母细胞表面塌陷的微绒毛又会再次树立起来,并且变得更加细长,此时的卵子最适合与精子接触融合。排卵后8 h还没与精子结合的卵母细胞开始老化,卵质膜上的微绒毛退化变粗,不利于精子的粘附和细胞膜的融合。
很早就有研究认为,ERM蛋白参与微绒毛的组装和维持,以确保其与胞质的强粘附性[43]。磷酸化的ERM蛋白通过连接肌动蛋白丝与整合膜蛋白形成微绒毛,常与微绒毛结构共定位。敲除ERM家族的radixin后卵母细胞表面微绒毛结构缺失,富集在微绒毛小泡中的卵母细胞分泌因子由于不能排出而堆积在卵母细胞中,抑制了颗粒细胞的增殖扩散,使卵泡发育水平整体降低,最终导致排卵异常,产仔率低,降低雌性哺乳动物的繁殖力[38]。小鼠卵母细胞中radixin的mRNA水平最高可达卵巢体细胞和其他组织中的700倍,其蛋白通常呈现激活状态,radixin在卵巢组织中存在如此大的组织特异性表达,可能是因为radixin在卵母细胞中承担着参与微绒毛形成的特殊功能[38]。
2.2 ERM蛋白调节卵母细胞的不对称分裂
与精子的产生不同,雌性哺乳动物的减数分裂最终只能形成一个成熟的卵母细胞。初级卵母细胞需要经过两次不对称分裂才能在形成单倍体的基础上保证大部分母源物质都留在卵母细胞中,以供早期胚胎的发育[44]。
卵母细胞的极性造成了纺锤体的不对称定位,从而导致了胞质的不均等分裂[45]。GVBD之后,卵母细胞纺锤体在胞质中心组装并向皮质区迁移,这一行为会导致覆盖于纺锤体上方的皮层形成一个富含肌动蛋白的区域,称为肌动蛋白帽[46]。这个区域几乎没有皮质颗粒存在,是极体最终排出的位置[47]。介导卵母细胞出现这种皮质区极化现象的是Ran-GTP信号通路[48]。染色体偏离中心向质膜移动使细胞出现了Ran-GTP浓度梯度,Ran-GTP浓度梯度通过CDC42激活皮质区的成核促进因子N-WASP,从而激活下游的肌动蛋白成核因子Arp2/3复合物,使纺锤体上方的皮质区积累大量肌动蛋白形成肌动蛋白帽[49]。同时肌动蛋白丝不断地从肌动蛋白帽流出,形成细胞环流,推动纺锤体向卵质膜移动[50]。到MII时期,纺锤体就会停留在皮质区附近,且平行于细胞皮层。一旦发生受精,卵母细胞被激活,纺锤体会发生旋转,改为垂直于细胞皮层,进而排出第二极体。但如果敲除ERM蛋白,卵母细胞的皮质张力会下降,纺锤体无法正常旋转,导致两极发生扭曲,向皮层顶起两个类似极体的凸起,第二极体不能排出,卵母细胞也不能向后发育[51]。
有研究发现,Ran-GTP酶也可以诱导ERM蛋白的去磷酸化,使肌动蛋白帽缺乏ERM蛋白活性,从而造成该区域没有微绒毛的覆盖(图4)[13]。微绒毛的区域性缺失也保障了卵母细胞的顺利分裂。因为微绒毛上富集着精卵结合蛋白,极体从微绒毛缺失的区域排出,可以避免精子从极体排出的位置入卵,造成父本遗传物质的丢失。这些研究结果都反映了ERM蛋白在维持卵母细胞的细胞极性,推动胞质不对称分裂等重要的生物学事件中扮演着重要角色。
3 ERM蛋白在精卵融合中的作用
精子与卵子的结合与融合是受精过程的关键步骤,依赖于精子和卵子之间复杂的分子识别体系和多对配体与受体间的协作,从而促使两个配子的质膜融合[52]。卵母细胞膜上的微绒毛是精子的初始结合点,可以捕获并包裹靠近卵母细胞的精子,将精子拽到卵子内部。该行为在受精过程里发挥着核心作用[53]。哺乳动物成熟卵母细胞表面是不均一的,面积较大的区域覆盖着密集的微绒毛,而减数第二次分裂中期纺锤体贴近的那小部分区域的卵质膜却是光滑的,这是因为精卵结合和融合的位点集中在微绒毛上,该区域缺乏微绒毛可以避免精子从纺锤体方向入卵使父系染色体随第二极体排出[54]。其实在很多体细胞中,微绒毛也同样担负着细胞粘附的功能,例如白细胞粘附分子L-选择素在微绒毛顶端表达,其尾部可以通过与ERM蛋白的N-末端结合连接肌动蛋白骨架,使白细胞粘附在内皮细胞上并沿着血管内皮滚动[55]。
CD9 (cluster of differentiation 9)是四跨膜蛋白超家族的成员之一,有研究发现该蛋白可以激活卵母细胞膜上的精子与卵子的融合位点;也可以借助外泌体的形式,将自身包裹在囊泡中从卵质膜释放到卵周隙,促进精子附着和精卵融合[56-59]。与磷酸化ERM蛋白的表达模式相似,CD9覆盖在卵母细胞的表面,但成熟卵母细胞不发生精卵融合的无微绒毛区域则没有CD9的表达[60]。卵母细胞敲除CD9后,微绒毛变得又短又粗[61],不易包裹精子,精子和卵子可以结合但最终无法融合,导致受精失败[62],注射外源CD9的mRNA后,受精率大幅度提高[63]。而敲除CD9基因的小鼠虽然可以正常排卵,但是精子却都被阻挡在卵周隙,无法与卵子融合,导致雌性的生育能力急剧降低[64]。免疫球蛋白超家族成员EWI-2和EWI-F是CD9的伙伴分子,其位于胞质的结构域中含有带正电荷的氨基酸,可以直接与ERM蛋白氨基末端的结构域结合,共同定位于细胞表面的微绒毛区域[65]。CD9与EWI-2和EWI-F结合形成复合体后,可以借助EWI-2和EWI-F与ERM的互作进而与微绒毛核心的肌动蛋白丝偶联在一起。也就是说,虽然CD9蛋白需要与细胞膜上的整合素等膜受体相互作用共同调控受精过程,但是只有通过CD9-EWI-2/EWI-F-ERM复合体将卵母细胞表面的精卵融合蛋白间接锚定到细胞骨架上,才能调节最适合精卵融合的微绒毛的动态结构,保障这些蛋白正常发挥作用[57]。
糖基磷脂酰肌醇锚定蛋白(GPI-APs)是一类借助自身羧基末端糖基-磷脂酰肌醇基团锚定到细胞膜外侧的蛋白质,不含跨膜组分[66]。磷脂酰肌醇磷酸脂酶C(PI-PLC)可水解大多数GPI-APs。用PI-PLC剥离卵质膜上的GPI-APs,精卵可以结合但不能融合[67]。叶酸受体4(FOLR4)也是一种参与精卵融合的GPI锚定蛋白,位于卵质膜上,与精子膜上的IZUMO1是一对精卵融合蛋白,所以又被命名为IZUMO1R或JUNO,这两种蛋白是精子和卵子能够认出对方的“身份证”。JUNO在成熟的卵母细胞上高表达,精卵结融合后JUNO会通过微绒毛上的小泡释放到膜外,受精40 min后就已经检测不到了,有研究认为JUNO受精后的脱落机制可能是为了防止多精受精[68]。两种膜蛋白的结合也需要CD9的协助,比如CD9会在受精前将JUNO招募到精卵融合位点,促进JUNO与IZUMO1的结合[69]。修饰后的GPI-APs到达内质网出口位点(ERES)由细胞质被膜复合体Ⅱ(COPⅡ)形成分泌小泡后从内质网转运到高尔基体,再运输到细胞膜表面,作为膜蛋白发挥功能[70]。有研究发现,ERM家族Radixin和Moesin可能通过介导细胞骨架重组参与驱动COPⅡ囊泡的运输从而影响GPI-APs的转运[71]。
4 ERM蛋白阻止多精受精
胎盘类哺乳动物是单精受精动物,只有一个精子与卵子结合才能保证形成二倍体受精卵。多精受精会使囊胚率降低,流产和畸形胎概率升高[72]。未成熟或老化的卵母细胞都容易多精受精,所以受精时就要求卵母细胞处于良好的胞质成熟状态。
精子入卵后,迅速激起卵质膜下皮质颗粒的大量胞吐,该皮质反应现象是卵子阻止多精受精的重要机制。例如,小鼠卵母细胞单精注射诱发皮质反应,处理后的卵子进行体外受精会发生多精入卵。皮质颗粒是卵母细胞特有的细胞器,但目前关于它的形成机制还存在争议,有研究认为它是由高尔基复合体分泌[73],也有相反的主张,认为皮质颗粒与滑面内质网小泡密切相关[74],但对于皮质颗粒在阻挡多精入卵方面的重要性却是得到研究者广泛认可的。随着卵母细胞的生长和成熟,皮质颗粒开始形成并且数量不断增加。在啮齿类动物中,皮质颗粒的重新分布有明显的极性,定位与微绒毛相似,与微丝帽的分布互补,成熟卵母细胞覆盖纺锤体的皮层没有皮质颗粒。而牛、猪和人等物种的MⅡ期卵母细胞中,不能看到明显的缺失皮质颗粒的区域,有研究发现这是因为该区域太小,镜下不能清楚观察到[75]。未成熟卵母细胞由于皮质颗粒数目不足且尚未贴近卵质膜,不能形成完善的皮质反应来阻止多精入卵[76]。“过熟”卵子中的皮质颗粒会内迁或提前胞吐,受精时皮质颗粒不能完全释放到卵周隙,容易造成多精入卵[77]。
皮质反应阻挡多精受精的原理是非常复杂的。精子先被透明带的ZP3糖蛋白识别,诱发顶体反应后与ZP2糖蛋白结合才能到达卵质膜与卵子融合[78]。精子的磷脂酶C(PLC)水解卵质膜上的PI(4,5)P2,生成二酰基甘油(DAG)和1,4,5-三磷酸肌醇(IP3)两种第二信使分子;IP3结合内质网上的IP3受体促进卵子内Ca2+释放并激活钙调蛋白依赖的蛋白激酶Ⅱ(CaMKⅡ),DAG则负责激活蛋白激酶C(PKC),使肌动蛋白结合蛋白MARCKS磷酸化,最终钙调蛋白与MARCKS结合,重塑肌动蛋白网络,使皮质颗粒发生胞吐[79]。
目前已经发现两条皮质反应阻挡多精受精的途径。首先,经皮质反应释放到卵周隙的皮质颗粒内容物中富含蛋白水解酶、卵过氧化物酶、N-乙酰氨基葡萄糖苷酶、糖基化物质等[28]。这些物质会改变透明带的结构,使其阻止其余精子入卵[80]。例如,SAS1B也叫ovastacin,是一种卵巢组织特异性表达的限制性金属内切蛋白酶,也是皮质颗粒的一种成分,最初分布在GV期卵母细胞的胞质中,随着卵母细胞的发育,SAS1B向边缘移动,最终定位于MⅡ的微绒毛膜上[81],受精时与精子顶体膜的SLLP1结合,使两层膜接触[82]。受精后皮质颗粒发生胞吐,SAS1B的表达量逐渐下降,到囊胚期几乎检测不到。精子入卵后,SAS1B随皮质颗粒排出,将透明带上的ZP2蛋白裂解,使透明带不能与精子结合,阻止多精受精[83]。同样是通过透明带反应阻止多精受精,卵母细胞与ERM蛋白家族中的ezrin蛋白共同孵育后,会增加透明带的消化时间,减少透明带上结合的精子数量,从而降低多精子入卵率[84]。并且,有研究发现精浆中也含有ezrin蛋白,会在第一个精子入卵后阻滞其余精子进入卵母细胞[85]。同时,精子膜、皮质颗粒膜和卵质膜都发生了复杂的融合,可能改变了卵质膜的功能元件,从而在卵质膜水平上阻止多精受精[86]。不同的物种会采用不同的方法阻挡多精入卵。例如,狗、山羊受精卵的卵周隙几乎没有精子,证明这些物种主要依靠透明带反应阻止多精入卵;兔、猪受精卵的卵周隙存在大量精子,表明他们对多精受精的阻止主要在卵质膜水平上;小鼠卵周隙存在少量精子,证明小鼠可以利用两种途径来保障单精入卵。有研究发现,ERM蛋白家族与肌动蛋白、肌球蛋白-II共同介导卵母细胞的细胞力学,通过微丝重排牵引皮质颗粒由胞质向细胞皮层移动,到卵母细胞成熟时皮质颗粒会沿质膜呈线状单层排列,这是皮质颗粒行使一切功能的基础[87]。
5 问题与展望
ERM家族特有的蛋白结构让它成为使很多膜蛋白与肌动蛋白形成互作关系的重要介质。因此,ERM蛋白能够作为肌动蛋白骨架与膜蛋白的交联剂参与调节减数分裂极体的排出、卵母细胞与颗粒细胞间微绒毛的动态组装以及成熟后的精卵融合等关键细胞活动。虽然ERM蛋白决定着卵母细胞的成熟质量和受精卵能否正常形成。但是多年来很多研究者大都着眼于ERM蛋白在白细胞迁移和各类癌细胞增殖等病理领域的功能探索,对ERM蛋白在哺乳动物繁殖层面的研究尚有不足。近年来的文献也只是浅显地提到了ERM蛋白会影响卵母细胞减数分裂过程和受精指数,缺乏对该蛋白作用机制的深入挖掘。未来从分子机制和生理水平上深入研究ERM蛋白的功能可能会是提高卵母细胞体外培养成熟率、保障后续的受精卵形成和胚胎发育的关键。



