刘传霞,王 晓,李雪雯,鲍苗菲,李婷婷,陈 欣,翁长江,郑 君
(中国农业科学院哈尔滨兽医研究所 动物疫病预防控制国家重点实验室 国家非洲猪瘟专业实验室 基础免疫创新团队,哈尔滨 150069)
非洲猪瘟(African swine fever,ASF)是一种具有高度传染性的病毒性疾病,可以导致家猪的高感染率和死亡率,但在自然猪群宿主[1-3]中表现为无症状。目前由于缺乏有效的疫苗和防治药物,该病已经造成了全球生猪产业的重大经济损失。现有的疾病控制方法是对受影响的地区进行检疫和扑杀被感染的动物。
ASF的病原是非洲猪瘟病毒(African swine fever virus,ASFV),是一种有囊膜的双链DNA病毒,可引起家猪全身出血性疾病[4]。ASFV是非洲猪瘟病毒科非洲猪瘟病毒属的唯一成员,是目前仅有的虫媒DNA病毒。病毒基因组为线性双链DNA,包括中间的稳定区和两端的可变区,大小为170~190 kbp,编码150~167个开放阅读框。ASFV病毒粒子为二十面体的多层结构,直径约250 nm,由外到内依次是病毒外囊膜、衣壳、内膜、内核心壳和病毒基因组[5]。ASFV编码的蛋白约200种[6],其中超过50种为结构蛋白[7],行使多种功能,包括病毒粒子形态的维持,吸附入侵宿主细胞,影响宿主细胞多种因子的功能以及调控宿主细胞的自噬、凋亡和炎症反应等防御机制[8]。
pE120R是ASFV编码的病毒衣壳蛋白之一,是一种晚期结构蛋白,由120个氨基酸组成[8-9]。pE120R主要定位于细胞质中,参与了ASFV从组装位点到质膜的运输,这表明pE120R在病毒的传播过程中起着重要作用[10]。免疫电镜观察显示,pE120R蛋白定位于细胞内病毒粒子的表面,以独立的方式与病毒DNA和主要结构蛋白p72相互作用,并且可能在病毒DNA的包封过程中发挥作用[11]。
1 材料与方法
1.1 细胞、质粒、实验动物及病毒株
E.coliDH5α、E.coliBL21(DE3)、HEK293 T细胞、pGEX-6p1(+)、pCAGGS-Flag-pE120R均由本实验室保存;SP2/0细胞由本实验室保存;猪肺泡巨噬细胞(PAMs)分离自30日龄ASFV阴性猪;BALB/c小鼠购自辽宁长生实验动物技术有限公司;ASFV HLJ/18株由中国农业科学院哈尔滨兽医研究所分离并保存。
1.2 主要试剂
DMEM、PBS、胰酶购自哈尔滨兽医研究所动物卫生中心;BamH I/NotI限制性内切酶、PrimeSTARMax Premix购自TaKaRa公司;弗氏不完全佐剂购自Sigma公司;同源重组试剂盒购自Vazyme公司;红外荧光标记的山羊抗兔和鼠的IgG购自LICOR公司;小鼠单抗Ig类/亚类鉴定用ELISA试剂盒购自博奥龙公司。
1.3 主要仪器设备
ELISA酶标板购自Greiner公司;细胞培养箱;高速离心机;ZEISS荧光显微镜;SDS-PAGE电泳仪及罗氏转印槽;Bio-Tek公司酶标仪。
1.4 pE120R基因重组质粒的构建
根据GenBank中登录的ASFV HLJ/2018株基因组组序列,设计引物pE120R-F: 5′-CAGGGGCCCCTGGGATCCATGGCAGATTTTAATTCT-3′(下划线处为BamH I酶切位点)pE120R-R: 5′-CAGTCACGATGCGGCCGCTTACTTCGATTT-ATGCGA-3′(下划线处为NotI酶切位点),由吉林省库美生物科技有限公司合成。以pCAGGS-Flag-pE120R为模板PCR扩增E120R基因。反应体系为50 μL:包括PrimeSTAR Max Premix(2×)25 μL,上、下游引物各1 μL,DNA 模板50 ng,ddH2O补至50 μL。PCR反应条件:98 ℃预变性 2 min;98 ℃变性10 s,55 ℃退火15 s,72 ℃延伸20 s,共35个循环;72 ℃延伸5 min;同时,pGEX-6p1(+)空载体经BamH I和NotI双酶切,将 PCR 产物和酶切产物经琼脂糖凝胶电泳鉴定后,用胶回收试剂盒纯化,然后二者经同源重组酶连接。连接产物转化DH5α感受态细胞,并涂布于氨苄抗性LB平板,37 ℃倒置过夜培养12 h。菌液PCR鉴定正确后,送至吉林省库美生物科技有限公司测序。测序正确的重组质粒按照小提质粒试剂盒说明书提取,命名为pGEX-6p1-pE120R,-20 ℃保存。
1.5 pE120R 重组蛋白的原核表达、鉴定与纯化
将表达质粒pGEX-6p1-pE120R转化大肠杆菌BL21(DE3)感受态细胞,挑取单菌落接种于5 mL氨苄抗性的LB液体培养基中,活化后加至2 L氨苄抗性的LB液体培养基,37 ℃,220 r·min-1培养OD600 nm至0.6~0.8时,加入终浓度为1 mol·L-1IPTG,在16 ℃,220 r·min-1培养16 h,离心,收集菌体沉淀。用PBS洗涤沉淀后再次重悬,冰浴条件下进行超声波破碎。收集上清液与GST Beads于4 ℃条件下孵育过夜,用不同浓度梯度咪唑缓冲液洗脱目的蛋白。然后对蛋白进行SDS-PAGE分析和Western blot鉴定分析,并进行蛋白定量,测定浓度后-80 ℃保存。
1.6 免疫小鼠
将纯化的pE120R蛋白进行浓度测定,将100 mg·mL-1的融合蛋白与等体积的弗式完全佐剂(首次免疫)和不完全佐剂混合并进行乳化,对5只BALB/c小鼠进行多位点皮下注射,免疫小鼠7周后断尾采血,离心分离血清,检测小鼠免疫后血清抗体水平,选择抗体水平最高的小鼠腹腔注射pE120R蛋白加强免疫,3 d后取该小鼠的脾进行细胞融合。
1.7 细胞融合及阳性杂交瘤细胞筛选
将加强免疫后的小鼠脾细胞与SP2/0 细胞进行融合,利用有限稀释法对筛选出来的阳性杂交瘤细胞进行3次亚克隆培养并通过间接ELISA筛选,以建立稳定分泌pE120R抗体的单克隆杂交瘤细胞株。对建株后的单克隆细胞进行扩大培养并冻存保种。
1.8 Western blot检测
将pCAGGS-Flag-pE120R重组质粒转染HEK293T细胞,24 h后收集细胞。同时利用1 MOI ASFV感染 PAMs,36 h收集细胞。用细胞裂解液充分裂解细胞,离心取上清,进行SDS-PAGE电泳并湿转至PVDF膜。将PVDF膜用含5%脱脂乳的TBS溶液室温封闭2 h,用pE120R单抗4 ℃作用过夜,洗涤,以红外荧光标记的山羊抗鼠IgG以1∶10 000稀释作为二抗,室温避光作用1 h,洗涤后用近红外荧光扫描成像仪扫描。
1.9 间接免疫荧光(IFA)检测
将pCAGGS-Flag-pE120R重组质粒转染HEK293T细胞,24 h收集样品。同时用1 MOI ASFV感染PAMs,分别于0、6、12、24、30 h收集样品,使用4%多聚甲醛常温固定pCAGGS-Flag-pE120R转染的HEK293T细胞和ASFV感染的PAMs细胞30 min,0.3% TritonX-100透膜10~15 min,5% BSA溶液封闭2 h,pE120R单抗4 ℃过夜孵育;添加相应二抗在室温避光孵育1 h。DAPI 室温染色15 min,置于共聚焦荧光显微镜下观察。
1.10 抗原表位的鉴定
分别以提取的ASFV HLJ/18株基因组DNA为模板扩增相应pE120R全长和各截断片段并克隆至pET-21a-MBP载体中,构建的重组质粒均测序鉴定正确。将上述重组质粒分别转化到BL21(DE3)感受态中,经IPTG诱导表达后以单克隆抗体和鼠源His抗体为一抗,以IRDye 800CW标记的山羊抗小鼠IgG(1∶10 000)为二抗,利用Western blot鉴定表位。
1.11 抗体亚型鉴定
取细胞上清50 μL用标本稀释液稀释(1∶1),加入到酶标微孔板中,同时设置阴性对照和阳性对照,37 ℃孵育30 min;弃去板内液体,用清洗液洗5遍,加入100 μL酶标二抗,37 ℃孵育30 min;弃去板内液体,用清洗液洗5遍,加入显色剂A和显色液B各50 μL,避光30 ℃孵育20 min,;加入终止液50 μL终止反应后用酶标仪测定450 nm处波长。
2 结 果
2.1 pE120R重组原核表达质粒的构建
首先利用TMHMM软件对pE120R蛋白的氨基酸序列进行分析,结果显示pE120R是一个无跨膜区的蛋白(图 1A)。根据E120R基因的编码序列(369 bp)设计引物,扩增E120R片段,PCR产物克隆至pGEX-6p1 载体。结果显示,PCR扩增条带单一,大小约369 bp,BamH I和NotI双酶切重组质粒,可见一条大小约369 bp的条带(图 1B),与预期结果一致。测序结果进一步证实该片段与pE120R编码序列一致,说明重组质粒构建成功,命名为pGEX-6p1-pE120R。
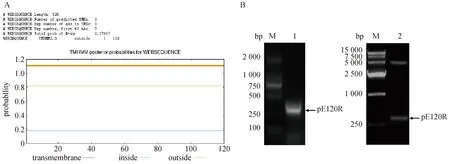
A. TMHMM 软件分析pE120R 蛋白的氨基酸序列;B. ASFV pE120R基因重组原核表达质粒的构建:M. DNA分子量标准(DL2 000,DL15 000);1. ASFV pE120R 基因PCR产物;2. pGEX-6p1-pE120R重组质粒双酶切鉴定 A. Analysis of amino acid sequence of ASFV pE120R protein by TMHMM software; B. Construction of the recombinant prokaryotic expression plasmid pGEX-6p1-pE120R: M. DNA Marker (DL2 000; DL15 000); 1. PCR production of ASFV pE120R gene; 2. Identification of pET-21a-pE120R plasmid digested with BamH I and Not I图1 重组原核表达质粒 pGEX-6p1-pE120R的构建Fig.1 Construction of recombinant prokaryotic expression plasmid pGEX-6p1-pE120R
2.2 pE120R 重组蛋白的原核表达、鉴定与纯化
SDS-PAGE结果显示,IPTG浓度为1 mol·L-1时,在16 ℃诱导16 h后,含pGEX-6p1-pE120R质粒重组菌样品中均出现了一条明显的蛋白条带,大小约25 ku,与预期大小相符,未诱导的含pGEX-6p1-pE120R重组菌液样品中未出现对应条带。SDS-PAGE和Western blot结果表明,pE120R蛋白以可溶性表达为主。经GST Beads纯化后获得了纯度较高的pE120R蛋白(图2)。
2.3 pE120R单抗隆抗体的Western blot检测
pCAGGS-Flag-pE120R质粒转染HEK293T细胞,24 h收集细胞。同时将ASFV感染PAMs,36 h后收集细胞,用含1% NP-40细胞裂解液裂解。以抗pE120R蛋白的单抗隆抗体为一抗,红外荧光标记的山羊抗鼠IgG(1∶10 000稀释)为二抗进行Western blot检测。结果显示:鼠抗pE120R蛋白单抗隆抗体可特异性识别外源表达的Flag-pE120R蛋白(图3A),以及ASFV感染PAMs后表达的内源性pE120R蛋白(图3B)。这些结果表明,制备的鼠抗pE120R蛋白单抗隆抗体可用于Western blot检测。
2.4 pE120R单克隆抗体的IFA检测
用pCAGGS-Flag-E120R质粒转染HEK293T细胞;同时将ASFV感染PAMs,经固定、透膜、封闭处理后,以抗pE120R蛋白的单抗隆抗体为一抗,将绿色(Alexa Fluor® 488)和红色(Alexa Fluor® 594)荧光标记的山羊抗鼠IgG以1∶2 000稀释为二抗。结果显示:鼠抗pE120R蛋白单抗隆抗体可以检测到外源表达的Flag-pE120R蛋白(图4A)以及ASFV感染PAMs后表达的内源性pE120R蛋白(图4B)。这些结果表明,制备的鼠抗pE120R单抗隆抗体可用于IFA检测。
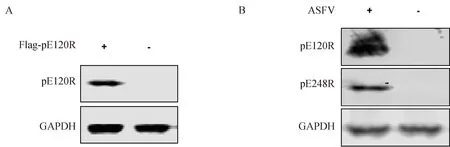
A. 鼠抗pE120R单克隆抗体Western blot检测Flag-tagged pE120R蛋白的表达; B. 鼠抗pE120R单克隆抗体Western blot检测ASFV感染PAMs pE120R蛋白表达 A. Detection of Flag-tagged pE120R by Western blot with mouse anti-ASFV pE120R mAb; B. Detection of ASFV pE120R protein expressed in ASFV-infected PAMs by Western blot with mouse anti-ASFV pE120R mAb图3 Western blot检测pE120R蛋白的表达Fig.3 Detection of pE120R protein expression by Western blot
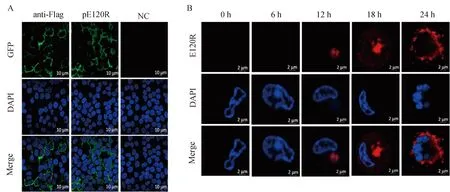
A. 鼠抗pE120R单克隆抗体IFA检测Flag-pE120R蛋白; B.IFA检测ASFV感染PAMs pE120R蛋白 A. Detection of pE120R by IFA with mouse anti-ASFV pE120R mAb; B. Detection of ASFV pE120R protein expressed in ASFV-infected PAMs by IFA图4 IFA方法鉴定Flag-pE120R 和 ASFV pE120R蛋白Fig.4 Identification of Flag-tagged pE120R and ASFV pE120R by IFA
2.5 抗原表位的鉴定
为确定抗原表位,将pE120R截短为6段进行原核表达,通过Western blot对截短进行鉴定。结果显示,该单抗的结合位点在30~60aa区域(图5)。
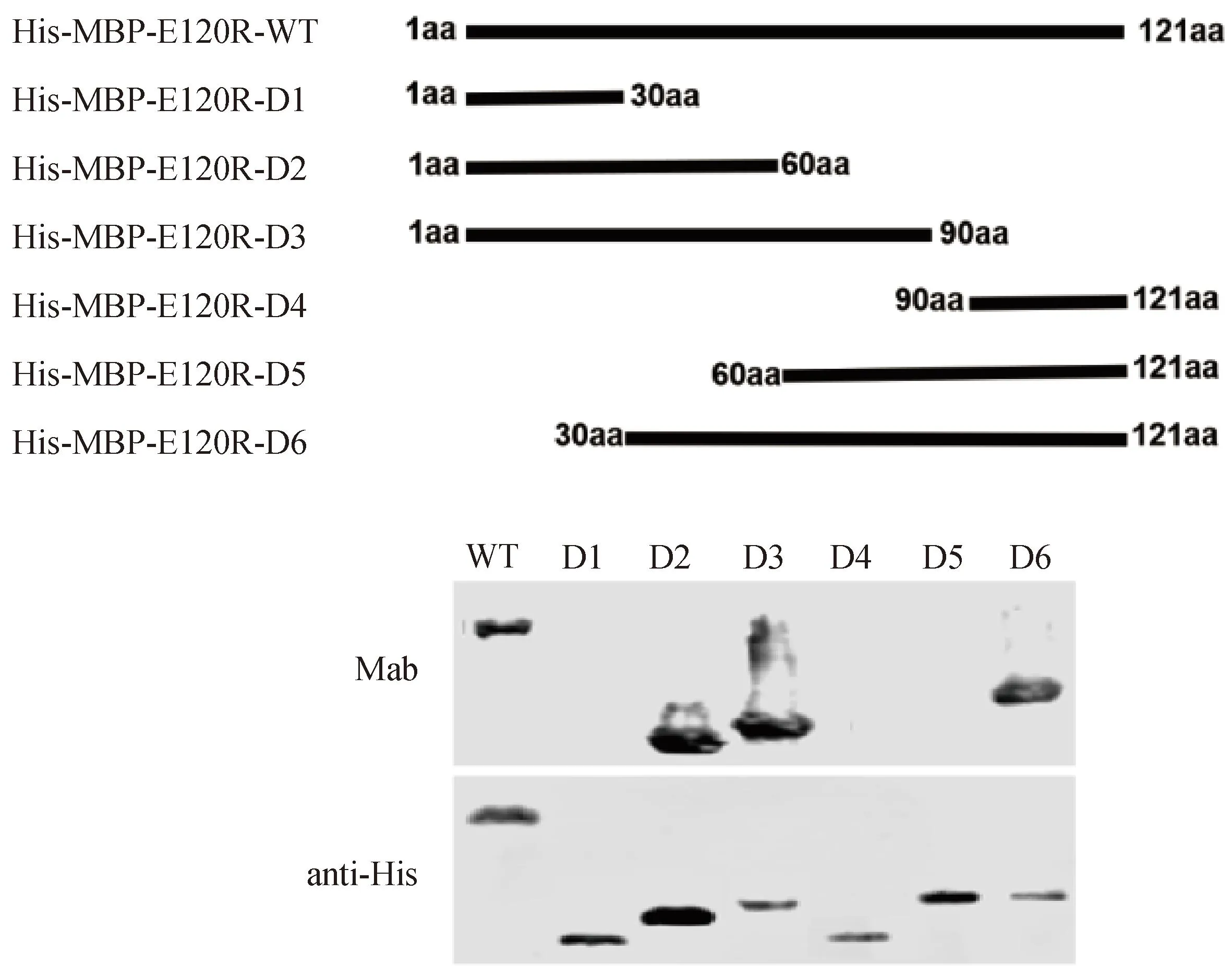
图5 Western blot鉴定单克隆抗体识别的抗原表位Fig.5 Identification of monoclonal antibody epitopes by Western blot
2.6 抗体亚类鉴定
小鼠单抗Ig类/亚类鉴定用ELISA试剂盒鉴定结果显示(图6),单抗识别的重链为IgG2a。

图6 单克隆抗体亚类鉴定Fig.6 Identification of monoclonal antibody subspecies
3 讨 论
猪肉是世界上消费最广、产量最高的肉类产品,占全球肉类产量的三分之一以上。猪的传染病威胁着这种重要的高质量蛋白质来源,而养猪业的全球化促进了病原体的出现和传播。在这些病原体中,ASFV已成为养猪业的“杀手”之一。
ASFV病毒结构复杂,其编码的很多蛋白都与结构和形态的发生有关,其中就包括pE120R蛋白。pE120R定位于成熟病毒粒子的衣壳上,是一种与主要衣壳蛋白p72相关的衣壳成分。一般来说,病毒在从宿主细胞进入和退出过程中的运动依赖于细胞骨架[12-14],而细胞骨架通常在整个感染过程中进行重组,而pE120R可能参与了ASFV在微管的运输,所以pE120R对ASFV的胞内运输具有重要的作用。此外pE120R还具有结合DNA的特性,这就表明pE120R很可能在病毒DNA复制、DNA封装或核蛋白核心组装中发挥作用。有研究发现敲除pE120R蛋白的ASFV在猪骨髓巨噬细胞(BMDM)中无法被挽救[15],pE120R还在天然免疫方面具有作用,它能够破坏TBK1-IRF3轴抑制宿主抗病毒应答[16]。目前关于E120R的研究还较少,需进一步探究。
本研究建立了能够大量制备可溶性pE120R蛋白的原核表达体系,并获得了高纯度的pE120R蛋白,可用于ELISA检测方法的建立等,具有一定的潜在应用研究价值。Western blot和IFA的鉴定结果表明,制备的抗pE120R单克隆抗体能特异性识别HEK293 T细胞中瞬时表达的Flag-pE120R蛋白和ASFV感染PAMs细胞后表达的pE120R蛋白。该单抗的结合位点在30~60aa区域,并且该单抗亚类鉴定重链为IgG2a。上述试验证明,制备的pE120R单抗具有较强的特异性和亲和力,为鉴定与pE120R相互作用蛋白和深入研究pE120R蛋白的生物学功能奠定了基础。
4 结 论
本研究利用原核表达系统纯化pE120R蛋白,筛选到1株稳定的杂交瘤细胞株,有望为ASFV的应用研究和基础研究提供帮助。



