黄大明 钟其冏 汤菁伟 Jose L.Hierro
(1.清华大学生命学院生态学实验室,北京 100084;2.蒙大拿大学生态系统研究所,美国蒙大拿 59812)
0.引言
生化它感概念由捷克-奥地利植物学家 Hans Molisch在他的著作《The Influence of One Plant on Another:Allelopathy》中首次提出[84]。书中提到许多植物通过释放刺激性或抑制性化学物质到环境中影响其他植物的例子,相互(allelo)感知(pathy)意味着植物除直接资源利用,还能以其他机制互动[110],以往常把生化它感误解成负相互作用[10,25,39,40,42,53,58,74,86,88,105,107,108],在过去20年中,许多研究显示生化它感释放的生化物质有抑制、兴奋、复杂信号对其他植物产生正负影响。一些信号可以依据环境条件被诱导释放,在某些情况下甚至是可逆的[81],生化它感作用不仅具有毒性作用[18,91,101,106,111],还有纯化学的正、负、中性作用,生化它感还广泛存在于植物与微生物,和微生物之间的相互作用中[25,36,54,51]。
以往生化它感的研究:加利福尼亚沿海鼠尾草灌木、森林和弃耕地空间格局中的生化它感[86,88,107],它感物质研究[7,40,52,58,68,115]。外来物种入侵中的生化它感[55,126];群落中优势种、入侵种的生化它感作用[15]。生化它感强度与物种协同演化程度[103],生化它感种内变异强度与共存[65],生化它感促进本地植物种与非本地种之间协同演化[63],等等[25,32,42,86,120,122]。本文讨论生化它感对物种分布,它感作用条件和物种多样性保持的影响。
1.物种分布
1.1 生态尺度
G.Evelyn Hutchinson在生态位理论中提出的生物制约与非生物制约,奠定了物种分布的理论基础[49],生物因子的相互作用可以增加基本生态位[14]。生态位可以是物种出现的生态或生物地理空间[73],推动了对生物和非生物因子与物种分布关系的研究[27]。当发现加州灌丛植物分泌毒素抑制禾草生长,影响分布格局;祁连山高寒草甸的细叶亚菊、萼果香薷等抑制邻体生长时,很多学者开始研究生化它感[88]。
1.2 生物地理尺度
生化它感是植物的个体特征,在个体生物场(生态位)作用显著,由于群落存在异质性,生化它感在景观群落中呈斑块状,不影响地理尺度的植物分布[73]。
1.3 分布与生化它感
生化它感是造成chaparral灌丛和加州沿海鼠尾草矮树丛下没有草本植物的原因[88]。研究发现灌木的挥发性化合物能抑制草本幼苗的根系生长,这些毒素存在于幼苗上的露水中。毒素能在空中有效地传播到邻近植物,而没有被土壤微生物腐化、土壤吸附或干燥[24,79,87-89]。还有一些研究认为:同一灌木物种中栖息着小型哺乳动物和鸟类,采食灌木附近的草本植物,而当这些食草物种被移除时,草本植物依然会重新在裸区生长[7,45],不承认分布与它感有关[40,58],接下来就有了40年的生化它感与物种分布的研究[75,76,92,115]。近些年,灌木与邻体间的化学相互作用研究有了新的转折点:三齿蒿 (Artemisia tridentate)分泌挥发性毒素抑制邻体种子萌发,如被食草动物采食,则邻体植物会生长,否则邻体植物被抑制。大量研究工作支持生化它感有助于加州chaparral灌丛(见图1)和其他区域植物的分布[48,57,120-122]。
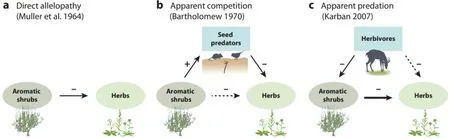
图1 在荒漠灌丛周围形成裸区的不同解释。(a)Mulle(1964)认为:灌丛通过产生挥发性毒素抑制邻体杂草和禾草[88]。(b)Bartholomew(1970)认为:灌丛为小哺乳动物和鸟类提供隐蔽场所,有利于采食附近的种子[7]。(c)为研究灌丛对邻体植物分布的生化它感作用,Karban(2007)发现灌丛的挥发性化合物会抑制邻体草本种子的萌发,且修剪植物会增加抑制剂的生产。实线和虚线分别表示直接和间接相互作用。线条的粗细表示作用的强度[56]。
入侵的禾本科高粱属草本植物(Sorghum)有很强的生化它感机制,能影响草地植被的格局。研究表明,双色高粱的细根会储存和释放大量的化学物质 sorgoleone,其他植物吸收后会干扰光合作用[29],入侵物种Sorghum halepense产生Sorgoleone[28]。强烈影响其他物种的分布。将S.halepense枝条和根部渗出液注入土壤会强烈抑制北美小须芒草 (Schizachyrium scoparium)的生长和繁殖[109]。同时,S.halepense的根茎每年扩张大约0.5m,降低S.scoparium在天然草地的多样性,直到最后移除。其他情况,入侵种如 Fallopia bohemica[94]和金合欢属(Acacia)[123]分别是由土壤生物群和其他植物化学助力。
根系生化它感是植物空间格局、形成植物领域性(防御和空间竞争)的基础[111]。北美荒漠Larrea tridentata灌丛的研究:1分布区内,成熟的L.tridentata灌木间隔非常规律[35]。2 L.tridentata 根部呈现离散空间分布,在不受制约的生长情况下,根冠直径比主茎要大4~5倍[12]。移除周围同种邻体,没有表现对水的竞争,不同个体的根之间几乎没有重叠[12,35,77],当同种根接近时,L.tridentata的根生长急剧下降。而在实验土壤中添加活性炭可以降低这种影响。表明:生化它感使根系离散,形成规则的空间分布格局[75,76],这种作用在共生植物Ambrosia dumosa也很明显,演替过程中,它几乎被L.tridentata竞争排除干净[78]。A.dumosa呈丛状分布[35],具有强烈不受活性炭影响的种内根-根接触,表现出适合一定尺度聚集而排外的生化它感[84]。L.tridentata,A.dumosa都没有表现出短期的地上种内竞争,主要是地下的资源分割[35]。类似的研究还有Glechoma hederacea(回避)和Fragaria vesca(入侵)的根反应,生化它感分别使其根部聚集和分散分布[31,113]。
2.物种相互作用的条件性
2.1 生态尺度
物种相互作用在强度和方向上随时空变化[13],在经典的拟谷盗(Tribolium)研究中,竞争结果受温度制约[95],土壤中磷含量制约着从互利共生到寄生的强度[44]。移除食植动物,植物间竞争会增强[17]。胁迫梯度假说 (SGH)认为:竞争与它感作用随环境中生物和非生物因子的胁迫而变化[11],机理简单,但强胁迫会产生次生物质[112],非生物因子可调节植物间生化它感强度[104]。
2.2 生物地理尺度
物种相互作用的条件会在地理时空范围发挥作用[13],在演化过程中,相互作用的物种会相互适应彼此变化,即协同演化。由于协同演化发生在相互作用物种的表型性状之间[44],并且因性状表达随时空变化,所以协同演化的现象也会显示出地理差异并受环境影响。由此构成协同演化地理镶嵌理论(GMTC)[118],包括3个基本的演化假设:区域选择性镶嵌假设、协同演化热点假设和性状再混合假设。区域选择性镶嵌假设假定物种特征的相互选择取决于区域环境条件;也就是说存在基因型×基因型×环境的作用影响物种的适宜度。协同演化热点假设提出:相互作用物种中性状的相互选择仅存在于特定群落(协同演化热点),这些群落嵌入自然选择不相互影响的区域(共同演化冷点)。最后,性状再混合假设:在一个区域内,具有物种以热点或冷点方式协同演化为特征的群落在基因层面上相连接,区域群落的遗传结构受到该区域内突变、基因流、遗传漂变和局部灭绝的影响[见图2(a)]。生物地理学的协同演化是影响生化它感作用强度的主要生物因素[103,105,15,106,46],而GMTC也是一个研究生化它感的角度。
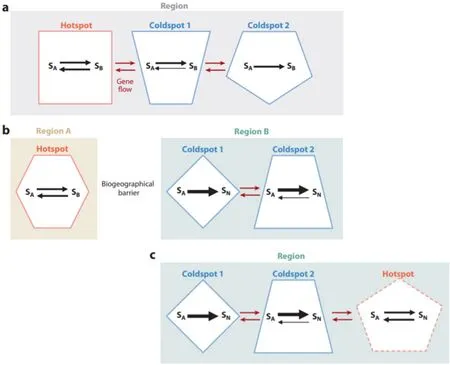
图2 (a)协同演化地理镶嵌理论(GMTC)示意图。协同演化镶嵌(由不同形状表示)发生在区域内并且在遗传上相互关联(红色箭头)。协同演化热点代表相互作用物种之间的性状的自然选择是互惠的区域,而协同演化冷点代表自然选择不是互惠的区域。区域内的黑色箭头表示:物种对彼此的选择,不同厚度代表不同选择强度;在五边形的冷点中,选择仅由物种A(SA)对物种B (SB)进行。(b)在GMTC背景下对物种入侵的生化它感研究。具有相同的演化历史的物种(区域A中的SA和SB)对彼此产生相似的相互选择现象,特别是在协同演化热点内,比没有相同演化历史的物种 [区域B中的SA和物种N(SN)]。SA起源于区域A并被引入区域B,在那里SN是本地种,并且对SA的植物化学物质敏感。(c)协同演化热点的形成。生化它感研究表明,随着时间变化,可能会在近期物种相互作用区域内出现协同演化热点(带虚线边框的镶嵌区域)。
2.3 生化它感的生态变异
生化它感会随着非生物因子的胁迫而增强,就是生化它感胁迫学说(SHA)[104,107],胁迫环境下,植物的生化它感物质会增加,次级化合物的生产也会增加[112,117]。许多研究显示:植物在生物和非生物因子的胁迫下都表现出增强生化它感[117,56,110],在干旱、半干旱和高海拔等环境胁迫中,生化它感作用会更频繁、更强烈,或者二者都有[97],而SGH则认为生化它感会增加正影响。由于它感产生的次级化合物和次级因素形成保护、营养和保水等效果超过抑制作用,而且还会在不同环境条件下,产生不同效果[4,80,93]。由于共生菌根帮助植物利用土壤资源,在胁迫环境中菌根尤为重要[67],会增强SHA[8]。
2.4 生化它感的生物地理变异
在生物地理尺度上,有协同演化史的物种间生化它感并不严重[15,62,103,104,105,106]。1974年俄罗斯生态学家Tikhon Rabotnov认为:生化它感作用在协同演化的植物群落(相同生物地理范围的物种间)中呈中性甚至微不足道,但在没有相同演化史的群落中很突出或严重[103]。后面的新型装备假说(NWH)[15]和生化识别假说(BRH)[106]都认为当物种遇到新生化信号时,会有高概率异常强烈的生化它感作用。
2.4.1 新型装备假说(Novel Weapons Hypothesis)
人类活动促进了外来物种入侵,使没有协同演化史的种群相遇,会产生协同演化冷点。也就是说,物种入侵可能会导致物种之间的选择不是相互的,而是强烈单向的[见图2(b)]。物种入侵和人造协同演化冷点是近来拉博特诺夫研究的主题,欧亚矢车菊(Centaurea diffusa)对北美物种的生化它感作用比对欧亚同系物种更强烈,这种差异在欧亚同系物种化学抑制磷吸收作用比北美同系物种强得多[15]。研究显示:一些非本地入侵物种会分泌发出对其原生群落中已适应的邻居相对无效的化学物质,可能是从营养利用途径,对侵入群落的植物具有高度抑制作用[71]。由此生化它感会导致外来入侵物种的丰盛度显著高于其原生区域[9,16,55,102,116,127],本地植物更容易受到外来植物代谢物的影响[126]。一般成功的入侵比不成功的入侵的研究要多,本地物种对非本地物种的相互抑制作用很少被关注。入侵不成功的例子:南美非本地草雀稗 (Paspalum notatum) 被一种原产于佛罗里达州的灌木Polygonella myriophylla的生化它感作用抑制[121];还有关于本地植物生化它感抑制非本地植物的研究[90,127]。此外,外来白羊草(Bothriochloa ischaemum) 的渗滤液和凋落物会降低本地大须芒草(Andropogon gerardii)和S.scoparium的萌发、生长和生存;而A.gerardii的渗滤液和凋落物还会降低B.ischaemum的生物质,而不影响S.scoparium[37],表明共同演化史对生化它感相互作用的强度很重要。
2.4.2 生化识别假说(Biochemical Recognition Hypothesis)
种子之间可以进行化学通信。种子可以通过识别同种或异种邻体释放的植物化学物质来分辨是竞争者[101,106]、促进者[123]或共生宿主[99],识别生物和非生物环境可以为萌发提供关键的信息,或诱导个体保持休眠状态直到有适宜的萌发条件。这是至关重要的识别机制,因为萌发是不可逆的,并且对适宜度影响深远,因此选择十分慎重[26]。具有相同演化史的种子会根据邻体情况调整萌发状态[106],通过研究阿根廷和北美草原共生物种的植物化学渗虑液发现:相对于异域物种,某些物种种子在接触过同域物种渗滤液后萌发率减少,但不影响幼苗生物量,表明种子存在物种特异性生化识别机制,只影响萌发,不影响生长,同域植物有利于物种特异性生化识别的演化发展[106]。用被非本地金合欢(Acacia)入侵和未入侵区域的渗滤液处理金合欢和本地其他物种的研究也有同样结果[123],但无论物种之间是否具有相同的演化史都会有一些促进萌发的表现。渗滤液对种子萌发的影响可能只是一种识别反应,而不是毒性作用。
2.4.3 共同演化热点的形成
在野外未曾遇过非本地臭椿 (Ailanthus altissima)的7个本地臭椿种更容易受到前者毒素的影响。这些种群产生的种子比遇过毒素的种群的种子更容易受到毒素的影响,表明植物对毒素的敏感性/耐受性是有遗传记忆的[69]。来自被矢车菊(Centaurea stoebe)入侵区域的一些物种表现出对C.stoebe的抵制性增强,且可以被活性炭改善[20]。相反,一些有相遇经验的草类对C.maculosa生物量竞争比没有相遇经验的同源草类更强。
伴随着适宜度优势和生产毒素成本之间的权衡,植物种可能会在非本地区域内减少或增加生化它感物质的生产。也就是说,如果植物生化它感代价过高,那么低它感个体会更受自然选择青睐,因为或许少量的毒素就足以抑制不适应毒素的邻体,当高浓度毒素对邻体植物无效时,释放低浓度毒素更有利于生存。另一方面,如果植物毒素可以提高生存优势,并且效果大于生产毒素的代价,增加毒素会更有利于自然选择,如同AARS假说的本地物种更具生化它感优势[19]。
有两项研究同时使用了本地和非本地基因型研究新环境中生化它感作用的演化发展。(1)欧洲本地基因型Alliaria petiolata以相同的比例减少了北美 Geum laciniatum和欧洲Geum Urbanum的萌发。然而,非本地北美 A.petiolata只减少了北美G.laciniatum的萌发,并对相遇过的欧洲G.urbanum没有影响[100]。原因是:A.petiolata的非本地种群沿着入侵时间序列响应环境竞争变化,在演化过程中减少了黑介子硫苷酸钾(即sinigrin,一种具有抗菌根特性的生化它感物质)的产量[61]。研究发现:随着非本地种群年龄的增长,群落越来越多地被A.petiolata主导,竞争从种间转移到种内。而在这种情况下,自然选择会降低sinigrin的产量,因为与群落中的其他物种相比,A.petiolata是非菌根的,而sinigrin的生产代价很高。由于上述工作的非本地种群采样可能来自A.petiolata繁茂的地方,因此sinigrin产量较低。低浓度的sinigrin可能对周边未相遇物种有效,但对周边相遇过的物种无效。但研究没有提到非本地种群与本地种群相比,sinigrin产量的变化。此外,A.petiolata对共生菌根的影响存在很大生物地理差异,使植物间的直接生化它感作用更复杂[16]。本地和非本地植物协同演化研究表明:与高sinigrin浓度非本地A.petiolata种群共同生长的本地Pilea pumila种群对A.petiolata的竞争适应性更高,反而对在低sinigrin浓度的P.pumila种群的竞争适应性更低[60,63]。(2)加拿大Solidago canadensis的非本地基因型,相对于本地基因型,增加了生化它感物质生产,并产生了增强抑制种子萌发和增加未相遇邻体生物量的物质,这些结果与AARS假说一致[125]。
结合对新型植物毒素的抗性增加[20,69]、本地物种竞争力的增强[20]、非本地物种的它感降低[61]、以及本地和非本地物种之间的相互作用[63],在没有相同演化史的区域,共同演化热点会频密出现[见图2(c)]。应该是物种共存的基础。
3.物种共存
3.1 物种共存理论
物种共存理论研究区域内生物和非生物因素对物种相互作用的影响,和在群落中的相对丰盛度[66,43],其核心就是探讨同一营养级物种的多样性保持机制。包含2个方面:均匀机制和稳定机制。均等机制减少了物种之间固有的适宜度差异,而稳定机制相对于反向的种间相互作用增加了反向的种内相互作用。也就是说:均匀机制使种内制约大于种间制约,而稳定机制与生态位差异有关[1,23]。二者的关键区别在于:稳定机制是丰盛度制约,均等机制则否[43],具有明显生化它感的群落是研究物种共存理论重要系统[1,66,70,74,114]。
3.2 非接触竞争与相互作用修改
大多数2个以上物种的研究都假定竞争关系具有等级性和接触性[114]。简单的3个物种,接触竞争可表示为物种A〉物种B〉物种C〈物种A,但也可以是非等级或非接触竞争,如物种A〉物种B〉物种C〉物种A,形成竞争环,没有优势种的群落构成石头剪刀布生态系统[59]。物种间共存不一定需要完整的竞争环才成立,重要的是:还有很多真正的循环未被认识[2,70,114]。特别是:一些物种在不形成竞争环的情况下影响其他物种间的相互作用,且保持强烈的间接相互作用。非接触竞争和形式竞争[46]对第三方物种相互作用的影响深受关注[46,82,70,114],网络格局的物种共存机制(即均等与稳定机制)还有待认识,生化它感、形式竞争等非接触作用在群落中的作用已有初步研究[46,60,61,2,82]。
3.3 共同演化
在探讨物种多样性保持机制时必须提到物种相互作用导致演化变化[66]。最适合用于解释在共存中演化重要性的例子是角色置换[30],即由竞争引起的生态位分化。演化不仅可以通过增加生态位差异促进物种共存,还可以增加物种之间适宜度相似性达到共存[66]。将演化的解释融合到Chesson框架的研究越来越多,而且生化它感在这些研究中扮演重要角色[47]。
3.4 非接触竞争的共存
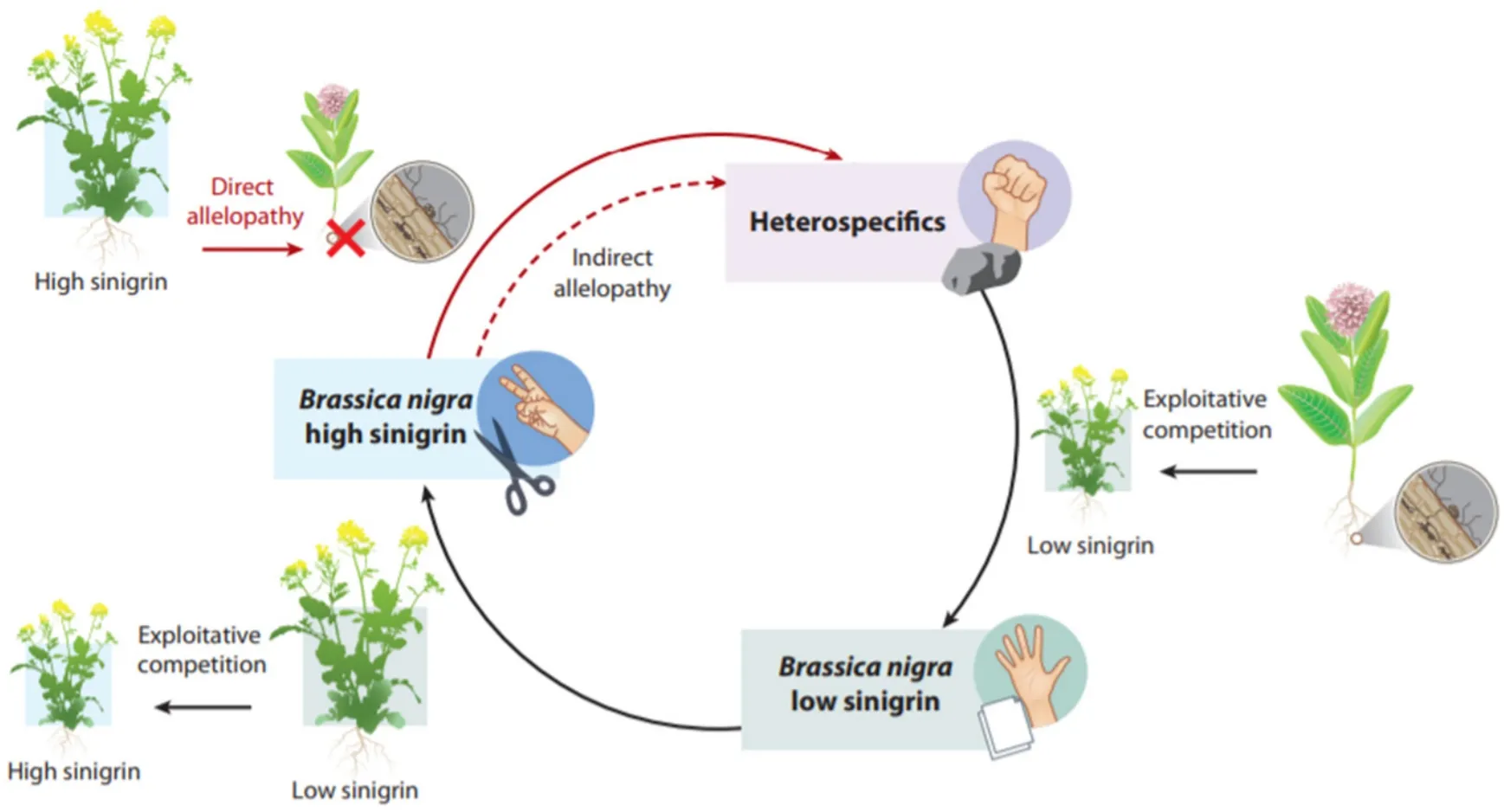
图3 黑芥(B.nigra)与邻体植物(异种)组成的非接触竞争格局。这种类型的循环动态也被描述成石头(异种)-布(低sinigrin浓度B.nigra)-剪刀(高sinigrin浓度B.nigra)系统。B.nigra 分别表达不同浓度的生化它感物质sinigrin;具有高 sinigrin 浓度的个体在竞争中排除异种,但被具有低 sinigrin 浓度的同种竞争,而后者又被异种排除。Sinigrin 对异种的抑制影响部分是通过抑制菌根实现。Sinigrin 生产成本高,而 B.nigra 是非菌根物种,因此需要在种内和种间竞争能力之间进行权衡。由于异种的菌根增强了它们对土壤资源的利用,得以抵抗低sinigrin浓度的B.nigra。这种竞争环保持了物种和基因型的多样性。该系统中物种的共存是由它感物的产生、群落中易感性和抗药性种群的存在、它感物生产和竞争能力之间的权衡、和基于资源和非资源作用的整合。以3种大肠杆菌菌株为代表的系统也有类似的结果。
群落中生化它感推动了早期的非接触竞争研究。最早的研究是珊瑚礁上固着无脊椎动物,而不是植物。海绵和海鞘对苔藓动物ectoproct特异性生化它感作用促进了竞争网络格局形成(即非接触竞争关系),从而阻止了任何单一物种独占海洋区域有限资源。尽管还没有理想的测量方法,但即使在没有干扰或捕食者存在的情况下,这种非接触生化它感可以保持高度的物种多样性[54],同样3种大肠杆菌菌株通过非接触生化它感达到共存。在这个体系中,大肠杆菌菌株C产生大肠杆菌素(colicin),一种可以杀灭易感性强的大肠杆菌菌株S的毒素,但大肠杆菌素对抗药性强的大肠杆菌菌株R无效。然而,菌株R的生长速度比菌株C快,因为生产大肠杆菌素比抵抗大肠杆菌素的成本更高,而且菌株S的生长速度比菌株R快,因为易感性提高了吸收营养的优势。因此,大肠杆菌菌株参与了上述的石头剪刀布对策[59]。重要的是,这项研究揭示了非接触竞争所需的条件:毒素、易感性和抗药性基因型的存在、毒素抗性优于毒素产生的成本平衡、以及所引起的不同类型负面影响(即生化它感和资源竞争[66])。由于天然植物、海洋固着无脊椎动物、真菌和细菌群落中都有可能存在相似的情况[59],非接触竞争是自然界中物种共存现象的重要机制[114]。而相比资源竞争,生化它感可能是重要的非接触因素[2]。
3.5 通过修改相互作用的共存
加利福尼亚稀树草原的 Quercus agrifolia 树下,一种本地草本植物Pholistoma auritum存在单种优势。在没有P.auritum的树下,树周围的一年生植物生长的很好,但P.auritum通过凋落物和渗滤液的影响能排除这些一年生植物[96]。在没有松树(Pinus ponderosa)冠层的草原上,入侵的杂草 Centaurea stoebe对入侵的草 Bromus tectorum具有很强的竞争性(可能是生化它感作用[108])。在单独生长环境下,即使松树凋落物释放的化学物质会影响其他入侵的物种,松树冠层对 C.stoebe 和 B.tectorum都没有影响,C.stoebe对B.tectorum 的竞争作用在 P.ponderosa下依旧没有发生。松荫也可降低 C.stoebe 的生化它感作用[22]。相似的研究还有P.ponderosa、C.stoebe与本地草Pseudoroegneria spicata之间的关系[82,83]。
3.6 共同演化与物种共存
生化它感的强度取决于种群共同的演化史的观点[15,103,105],说明同一营养级的物种之间的共同演化可能减少强烈不平衡相互作用的比例并促进共存[20,63],种群对邻体释放的化学物质的适宜度改变[34,20,38,69],以及它们之间的演化改变[63],可以促进物种共存[1,23,43]。研究 A.petiolata 和 P.pumila 之间共同演化关系中的共存机制[63]。发现稳定性机制(增加生态位差异)比均等机制(减少适应度差异)更能促进这些物种的共存。由于新型它感物质在非本地物种的优势大于本地物种[74]。强调通过稳定性共存机制(即生态位分化)可排除适宜度差异所导致的竞争排斥[1]。最近发现生化它感强度与物种系统发育之间存在负相关,表明生化它感可能有助于近亲物种的共存[47,126]。
3.7 通过化学识别的共存与超产中化学通讯作用
植物化学通讯非常复杂。它们能在地上和地下区别自身分泌物质和非自身分泌物质,近亲的和非近亲的。植物选择性识别和反应来自其他植物的化学信息能力会促进物种共存。
例如,松树(Pinus spicata)混合种群比单一种群生产力高(产量过剩,即超产)的现象,不能用资源互补或土壤病原体等传统机制来解释[5]。相反地,P.spicata 的种内产量过剩可能是收到根识别的影响[124]。也就是说P.spicata根部的伸展中在接触同种群的植物根比接触来自不同种群的植物根后下降更多。这意味着根系占据的土壤体积的减少,且单一种群中的资源获取比混合种群要多[124],关于根识别的研究很多[18,31,32,57,75,76,84,85,91]。
早期未把混合种群中的超产和化学相互作用联系起来。最近发现作物根系分泌物使间作系统中出现超产现象。在低磷土壤中生长时,蚕豆(Vicia faba)通过释放有机酸和氢离子使土壤酸化和激活磷离子,从而增加了玉米(Zea mays)的生物量和谷物产量。这些物质的释放促进了玉米对磷的吸收,正好玉米对磷有很高的要求[72]。反过来,玉米的根系分泌物增强了蚕豆根瘤的形成和氮的固定,提高了其生产力[71]。这些刺激作用是有特异性的,因为来自普通小麦(Triticum aestivum)和大麦(Hordeum vulgare)的根系分泌物不会促进蚕豆根瘤形成[71]。蚕豆-玉米间作体系的物种特异性现象很特别,因为在地理起源较远的物种之间(源于近东的蚕豆[21]和美洲的玉米[96])出现了相互促进的化学反应,并且从未在相似地理起源物种之间观察到(源于近东的小麦和大麦[41])。但不知道协同演化是否会影响正向化学作用,因为它会影响反向的生化它感[15]。野生植物的这种根部化学促进作用还没有研究。
4.结语
(1)最近研究显示:可以从生化它感角度认识植物间相互的抑制和促进作用。
(2)生化它感影响着世界各地草原、灌丛和森林中的区域植物分布。
(3)简单的增减物种实验可以证明一般的正/负相互作用会影响物种分布。但生化它感(或与此相关的营养稀缺)作为影响物种分布的机制需要积累相关的研究,而不是一个决定性的实验。也就是说,可以证明物种A抑制物种B,物种 A 产生一种毒性化学物质抑制物种B。但无法消除所有其他的潜在机制。
(4)生化它感的强度可能受共同演化历程制约,因此新型生化它感会促进外来物种入侵。
(5)本地物种对非本地它感物质的适应反应和非本地物种对本地它感物质的适应反应表明,共同演化热点可以在没有相同演化历程的物种相遇区域迅速出现。
(6)在已建立竞争环的远距离系统发育中,生化它感可以促进非接触竞争和物种共存。
(7)在基本生态学概念中,生化它感非常重要。
5.展望
(1)需要研究影响生化它感相互作用强度与方向的生态条件模式。
(2)通过沿胁迫整合生化它感,进一步探讨非生物环境因素影响物种相互作用。
(3)进一步关于大肠杆菌体系和黑芥体系共有的特征的研究,探讨非接触作用和物种多样性保持。
(4)本地和外来物种间生化它感与物种共存。
(5)对植物间正/反向化学识别机制研究,增加对生化它感的生态与进化认识。



