刘振春,郭 炫,杨童奥,梁国朋,吴 伟*
(吉林农业大学食品科学与工程学院,吉林 长春 130118)
填饲对鹅肥肝脂肪酸合成酶基因mRNA表达的影响
刘振春,郭 炫,杨童奥,梁国朋,吴 伟*
(吉林农业大学食品科学与工程学院,吉林 长春 130118)
以朗德鹅为对象,采用实时荧光定量PCR法研究不同填饲周龄脂肪酸合成酶基因(FAS基因)表达的规律性。结果显示,FAS基因相对表达量(FAS与β-actin基因表达量的比值)在填饲前、填饲1周、填饲2周、填饲3周和填饲4周相对表达量逐渐增高,分别为0.035、0.128、0.253、0.876、1.009,说明肥肝组织中FAS基因表达丰度与填饲期呈显着正相关,FAS基因在鹅肥肝形成过程及体脂沉积过程中具有一定的调控作用,为促进鹅肝内脂肪沉积的主要调控基因。
填饲;鹅肥肝;脂肪酸合成酶;基因表达
鹅肥肝主要是指朗德鹅生长发育基本完成后,在短期内(2~3周)人工对生长发育至一定体质量(一般为4.5kg左右)的鹅进行强制填喂过量玉米或糙米等高能量的碳水化合物饲料,使这些饲料通过鹅机体正常的新陈代谢,转化为甘油三酯(TG)并在肝脏内贮存而获得比正常鹅肝重5~10倍的脂肪肝[1],它是一种高档的新型家禽产品。鹅肥肝并不影响鹅的正常身体代谢,法国Grassey教授精辟地指出:“禽类吃了含多种糖分和大量淀粉的饲料后,生成沉积于皮下、腹腔和肝脏中的脂肪,这种脂肪的出现是一种正常状态,就像猪在皮下沉积脂肪是一个机理”。大量的脂肪沉积在鹅肥肝中,在质量和生理理化性状两方面都和超期饲养前鹅的普通肝有很大差异,因此鹅肥肝和鹅肝的概念是完全不同的。鹅肥肝含有蛋白质、脂肪、维生素、卵磷脂、甘油三酯、各种酶、核糖核酸、脱氧核糖核酸和多种微量元素等营养成分[2]。经填饲后的肥鹅肝,其营养成分发生了显着变化,脂肪含量高达60%~70%左右,是正常肝的7~12倍;不饱和脂肪酸比动物油中含脂肪较高的猪油还要高11%以上,比正常肝相对含量增加20倍;卵磷脂增加4倍;酶活性增加3倍;核糖核酸和脱氧核糖核酸增加1倍[3-4]。每100g肥肝中卵磷脂含量高达4.5~7g,脱氧核糖酸和核糖核酸含量达9~13.5g。
“卵磷脂”是当今国际市场保健药物和食品中必不可少的重要成分,具有降低血脂,软化血管,延缓衰老,防治心脑血管疾病发生的功效;而亚油酸为人体所必需,且在人体内不能合成,必须由食物中摄取,可降低人体血液中胆固醇的含量[5-6]。鹅肥肝与普通鹅肝相比,有效营养物质在体内氧化后产生的热量增加10倍[7],极大地提高了它的营养价值和食疗价值[8]。本研究利用实时荧光定量法测定鹅肥肝中脂肪酸合成酶基因(FAS基因) mRNA表达丰度变化规律,以验证FAS基因是鹅肥肝形成过程中关键调控基因,旨在为今后鹅肥肝填饲提供有价值的技术参数。
1 材料与方法
1.1 实验动物与样品采集
选取由吉林省9台顺达鹅业有限公司的同批孵化和在相同饲养管理条件下统一饲养的健康朗德鹅132只,体质量4.5kg,10周龄,并进行随机编号,填饲28d。每个填饲阶段结束后,随机抽取20只宰杀,迅速取肝脏10g左右,放入10mL冻存管中快速置入液氮中保存,带回实验室后尽快提取RNA,以免其降解。
1.2 试剂与仪器
焦碳酸二乙酯(DEPC) 美国Sigma公司;Trizol试剂 美国Invitrogen公司;氯仿 天津化学试剂有限公司;异丙醇 北京化工厂;反转录试剂盒 日本TaKaRa公司;DNA Marker DL2000 天为时代生物有限公司;DNA凝胶回收试剂盒 美国Axygen公司;小剂量快速质粒DNA提取试剂盒 美国Omega公司。
超净工作台 苏净集团安泰公司;核酸浓度测定仪 英国Pharmacia Biotech公司;ABI System 7000型荧光定量PCR仪 美国ABI公司;-80℃超低温冰箱 日本Sanyo公司;低温高速离心机 美国Sigma公司。
1.3 总RNA提取
采用Trizol一步法,用于总RNA提取的塑料器皿、耗材均经过0.1% DEPC处理,玻璃器皿于180℃干烤8h以上。具体操作步骤参照说明书。
将2μL RNA溶液加入198μL的0.1% DEPC处理的水中,以DEPC处理水为空白对照,用紫外分光光度计测定其在波长260nm处的光密度值(OD260nm)以及波长260nm与波长280nm处OD值的比值(OD260nm/OD280nm)及总RNA浓度。
1.4 反转录合成cDNA
采用AMV反转录酶,以提取的鹅肥肝总RNA为模板,以Oligo-dT为引物(工作浓度为10pmol/μL)进行反转录反应。
1.5 PCR扩增
根据GenBank中原鸡的FAS(NM_205155.1)和β-actin(L08165.1)的mRNA序列设计引物,由上海生工生物工程技术服务有限公司合成,详见表1。
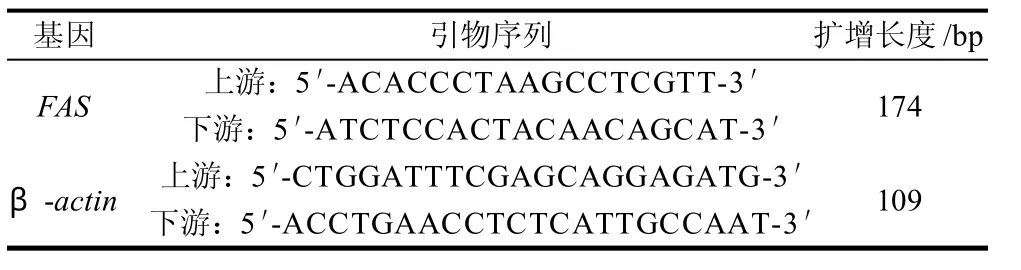
表1 RT-PCR反应所用引物Table 1 Primers of RT-PCR
运用常规PCR法,用内参引物以反转录产物为模板进行扩增,验证cDNA的完整性。然后扩增目的片段,得到的PCR产物可以用于制备荧光定量PCR的阳性对照标准品。
1.6 PCR回收
采用DNA凝胶回收试剂盒回收PCR产物目的片段,目的片段DNA溶液于-20℃保存。
1.7 目的片段克隆与测序
将凝胶回收的目的DNA片段连接到pMD-18T载体上,用CaCl2法制备E.coliDH5α感受态宿主菌(在严格的无菌条件下制备感受态),分别扩增128bp和316bp的目的片段和81bp的内参片段,将PCR鉴定正确的重组质粒送上海Sangon公司进行序列测定。
1.8 实时荧光定量PCR
将质粒溶液用纯水进行梯度稀释,分别稀释到1×109、1×108、1×107、1×106、1×105copies/μL,即为实验设计标准品的5个水平,每个水平设3个重复,3个阴性对照。加样过程中,要确保相同处理各管反应液的均一性,且确保各管加样量一致。另外还需注意防止96孔反应板上各管的交叉污染。
设置荧光定量PCR仪程序。与探针法不同的是,染料法需要附加一个溶解曲线制作反应。
2 结果与分析
2.1 鹅肥肝中总RNA的提取
所提取的RNA样品用分光光度计测定,所有的RNA样品OD260nm/OD280nm值均在1.8~2.1之间,总RNA电泳图有清晰的28S、18S条带(图1),表明所提取总RNA完整性较好,无降解,RNA质量符合RT-PCR实验要求。
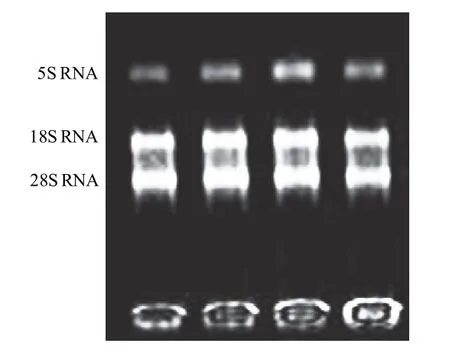
图1 鹅肥肝总RNA电泳结果Fig.1 Electrophoresis results of total RNA from tissues
2.2 实时荧光定量PCR检测的灵敏性
以梯度稀释的标准品为阳性模板进行实时荧光定量PCR反应得到的扩增曲线见图2、3。
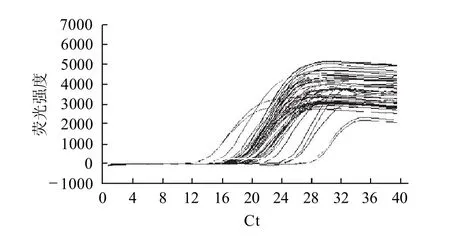
图2 FAS基因扩增的循环数与荧光强度的关系Fig.2 Correlation between cycle number of FAS gene amplification and fluorescence intensity
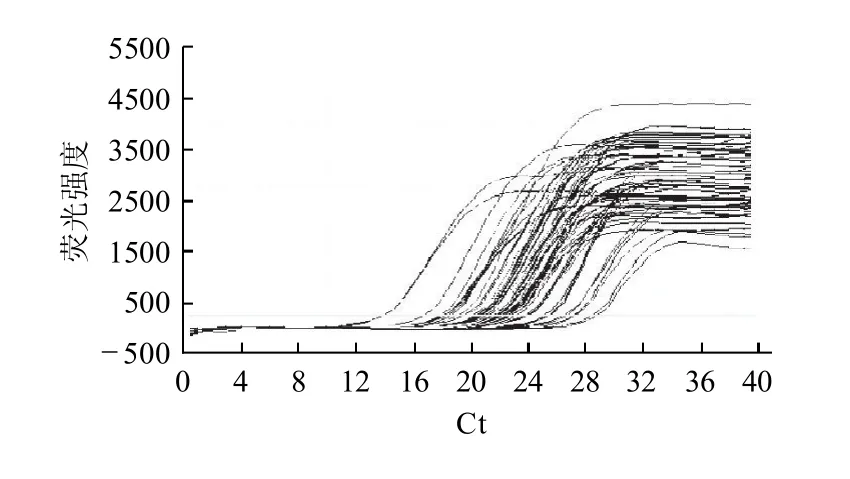
图3 β-actin基因扩增的循环数与荧光强度的关系Fig.3 Correlation between cycle number of β-actin gene amplification and fluorescence intensity
由图2、3可知,不同拷贝数的检测范围较广,循环阈值(Ct)范围在13~35,扩增前期曲线重合性好且无杂峰,呈典型S型荧光定量学曲线,是理想的扩增曲线,说明实时荧光定量PCR检测FAS基因具有较好的敏感性,反映出样品扩增效率较好。
2.3 实时荧光定量PCR扩增产物的溶解曲线和假阳性曲线
以不同拷贝数阳性标准品模板的对数为横坐标,以荧光定量PCR反应过程中出现荧光信号的初始Ct为纵坐标绘制FAS基因和β-actin基因的标准曲线,其中荧光信号的初始Ct是指图中不同拷贝数的模板与虚线相交处的循环数。FAS基因的标准曲线方程为:Y=-3.588x+14.447(R2=0.994),扩增效率为90.0%;β-actin基因的标准曲线方程为:Y=-3.684x+14.067(R2=0.994),扩增效率为86.8%;说明实验扩增效果好,具有较高的可靠性。
阳性模板在实时荧光定量PCR反应的后期,产物经熔融、冷却得到溶解曲线。其中横坐标代表荧光值(-d(RFU)/dT),纵坐标代表温度,见图4、5。
图4 FAS基因扩增溶解曲线Fig.4 Dissociation curve of FAS gene amplified product
图5 β-actin基因扩增产物溶解曲线Fig.5 Dissociation curve of β-actin gene amplification product
由图4、5可知,溶解曲线成单峰,说明扩增产物单一,避免了假阳性结果的出现,扩增产物特异性好。
2.4 填饲不同周龄FAS基因表达规律
采用相对定量,即FAS和β-actin荧光定量的比值(FAS/β-actin比值)来表示。FAS和β-actin荧光定量的比值测定结果见表2。填饲前、填饲不同周龄,鹅肥肝组织中FASmRNA表达量差异显着(P<0.05)。填饲前与填饲1周差异不显着(P>0.05),填饲3周、填饲4周与填饲1周FAS基因的mRNA相对表达量差异显着(P<0.05)。
由表3方差分析结果可以看出,不同填饲周龄鹅肥肝中FAS基因相对表达量逐渐增加。填饲前FAS基因相对表达量很低,仅为0.035±0.032,此时对应的肝质量为(105±31)g;填饲1周FAS基因相对表达量为0.128± 0.031,肝质量为(242±40)g,FAS基因相对表达量增加了0.093,肝质量增加了137g;填饲2周FAS基因相对表达量为0.253±0.049,肝质量(467±163)g,FAS基因相对表达量增加了0.125,肝质量增加了225g;填饲3周FAS基因相对表达量为0.876±0.162,肝质量(805± 181)g,FAS基因相对表达量增加了0.623,肝质量增加了338g;填饲4周FAS基因相对表达量为1.009±0.435,肝质量(995±220)g,FAS基因相对表达量增加了0.133,肝质量增加了190g。表明FAS基因相对表达量在填饲第3周时增加迅速,肝质量增加量也大,而填饲第4周,FAS基因相对表达量增加少,肝质量增加也随之减少。
根据表3结果,对FAS基因相对表达量(y)与填饲周龄(x)进行似合,得线性回归方程为:y=0.2696x-0.2138(R2=0.8994),说明FAS基因相对表达量与填饲不同周龄呈正相关,填饲周龄越长,FAS基因相对表达量越高,但后期(填饲4周)增加不显着。量不断增加,增加趋势与肝质量增加速度基本一致,特别在填饲第3周时FAS基因相对表达量增加达到最高值,此时肝质量增加也达最高值338g,鹅肥肝质量也超过750g,随后FAS基因相对表达量趋于平稳,肝质量增加缓慢,说明填饲前期(1~3周)脂肪主要沉积在肝内,填饲超过21d脂肪就会向肝外组织转移。同时随着填饲量和填饲次数的增加,FAS基因在填饲不同周龄的相对表达量也逐渐升高,脂肪在肝内外的沉积量也随之增加,从而表明填饲对基因FAS的表达影响及其对鹅肥肝增重的作用是非常显着的。但Nogalska等[9]研究表明,不同日龄猪腹脂FAS基因表达量随日龄增大呈逐渐增加的趋势。

表3 不同填饲期FAS基因的相对表达量测定结果Table 3 Expression of FAS/β-actin in different feeding periods
3.2FAS基因对鹅肥肝脂肪形成的调控作用
荧光定量PCR结果显示FAS基因相对表达量在填饲前,填饲1~4周逐渐增高,分别为0.035、0.128、0.253、0.876、1.009。FAS基因相对表达量与填饲期显着正相关(R2=0.8994,P=0.037<0.05)。Badger 等[10]研究发现,鸡肝FAS丰度越高越能显着提高体内TG的沉积,其活性与肝脏中脂肪的合成与体脂沉积呈正相关。刘涛[11]研究表明,绵羊尾部脂肪组织中的FAS基因表达丰度与体质量呈显着正相关。Reiter等[12-13]研究发现,猪肝脏组织中FASmRNA表达丰度与体脂肪含量呈正相关,这与本研究结果相一致,表明FAS基因是鹅肝脂肪沉积的主要调控基因,其表达量的高低直接影响脂肪在肝内外的沉积量。
3 讨论与结论
3.1 鹅肥肝组织中FASmRNA表达量与产肝性能的相关性分析
本研究得出,FAS基因相对表达量与鹅肥肝质量呈正相关。填饲前FAS基因表达量很低,鹅肝质量只有(105±31)g,随饲喂周龄不断延长,FAS基因相对表达
[1] 蔡云升. 鹅肥肝与鹅肥肝酱的开发[J]. 食品工业, 2003(6): 48-49.
[2] HERNIER D, ROUSSELOT-PAILLEY D, PERESSON R, et al. Influence of orotic acid and eserogen on hepatic lipid storage and secretion in the goose susceptible to liver steatosis[J]. Biochimica et Biophysica Acta, 1994, 1211(1): 97-106.
[3] YAMAMOTO A, ISOZAKI M, HIRAYAMA K, et al. Influence of dietary fatty acids on phospholipid fatty acid composition in subcellular particles of rat liver[J]. Journal of Lipid Research, 1965(6): 295-300.
[4] BELL M V, HENDERSON R J, SARGENT J R. The role of polyunsaturated fatty acidsin fish[J]. Comp BiochemPhysiol B, 1986, 83(4): 711-719.
[5] 瞿浩, 王继文. 鹅肥肝形成的分子机理研究进展[J]. 四川畜牧兽医, 2003(5): 33-34.
[6] BODI L. A study of liver production ability in hungarian and landes geese after plucking[J]. Allattenyersztes es Takarmanyozas, 1992, 41(2): 123-131.
[7] 王炜, 张伟敏. 单不饱和脂肪酸的功能特性[J]. 中国食物与营养, 2005 (4): 44-46.
[8] 葛长荣, 苏子峰, 曹正辉. 多不饱和脂肪酸免疫功能的研究进展[J].饲料工业, 2005, 26(8): 1-4.
[9] NOGALSKA A, SWIERCZYNSKI J. The age-related differences in obese and fatty acid synthase gene expression in white adipose tissue of rat[J]. Biochemical et Biophysica Acta, 2001, 1553: 73-80.
[10] BADGER A M, NEWMAN-TARR T M, SATTERFIELD J L, et al. Regulation of fatty acid synthase(FAS)[J]. Progress in Lipid Research, 1997, 36(1): 43-53.
[11] 刘涛. 不同能量水平日粮对绵阳脂肪酸基和肥胖基因表达的影响[D].兰州: 甘肃农业大学, 2007: 48-52.
[12] REITER S S, HALSEY C H C, STRONACH B M. Lipid metabolism related gene expression profiling in liver, skeletal muscle and adipose tissue in crossbred Duroc and Pietrain pigs[J]. Comparative Biochemistry and Physiology Part D Genomics Proteomics, 2007, 2 (3): 200-206.
[13] 单体中, 汪以真, 刘建新. 不同日龄猪腹脂中脂肪酸合成酶(FAS)基因表达规律的研究[J]. 畜牧兽医学报, 2006, 37(7): 662-666.
Effect of Feeding Period on Expression ofFASmRNA in Fatty Liver of Goose
LIU Zhen-chun,GUO Xuan,YANG Tong-ao,LIANG Guo-peng,WU Wei*
(College of Food Science and Engineering, Jilin Agricultural University, Changchun 130118, China)
Landes geese were used as the materials to explore the expression rule ofFASmRNA in fatty liver through real-time fluorescence quantitative PCR method during overfeeding weeks. Fluorescence quantitative PCR results showed that the expression ofFASgene revealed an obvious increase during pre-feeding fold and pro-feeding 1 week, feeding 2 weeks, feeding 3 weeks and feeding 4 weeks, which were 0.035, 0.128, 0.253, 0.876 fold and 1.009 fold enhancement, respectively. A positive genetic correlation betweenFASmRNA expression in fatty liver tissue and feeding weeks was observed. These results suggested thatFASgene had a certain readjustment function during the developing process of fatty liver in goose and the process of body fat deposition. Thus,FASis a regulation gene to promote fat deposit inside liver of goose, which provides valuable technical parameters for future studies of fatty liver.
feeding;fatty liver of goose;fatty acid synthase;gene expression
TS251.1
A
1002-6630(2012)05-0165-05
2011-04-07
吉林省科技厅科技发展计划重点项目(2009026)
刘振春(1963—),男,教授,博士,研究方向为食品营养与功能性食品。E-mail:liuzhenchun63@163.com
*通信作者:吴伟(1955—),男,教授,博士,研究方向为动物营养与种性。E-mail:jilinww@yahoo.com.cn



